

Properties and functional role of voltage-dependent potassium channels in dendrites of rat cerebellar Purkinje neurons
- PMID: 12843273
- PMCID: PMC6741279
- DOI: 10.1523/JNEUROSCI.23-13-05698.2003
Properties and functional role of voltage-dependent potassium channels in dendrites of rat cerebellar Purkinje neurons
Abstract
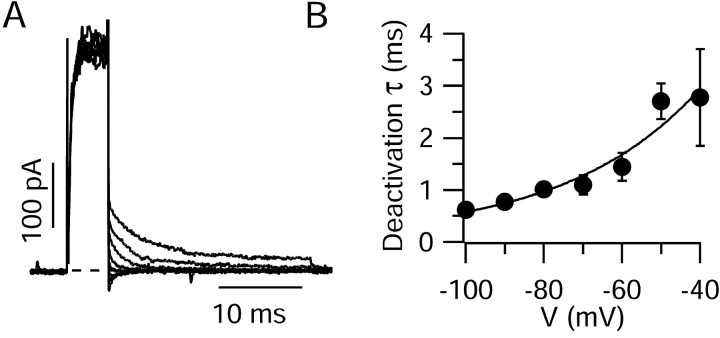
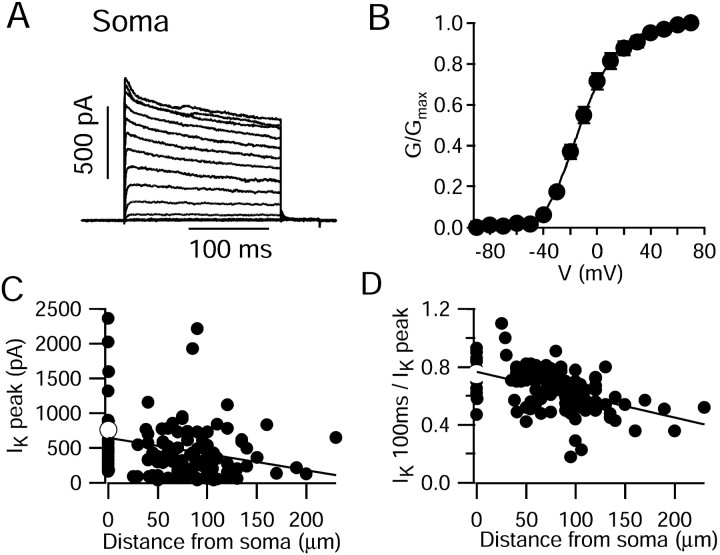
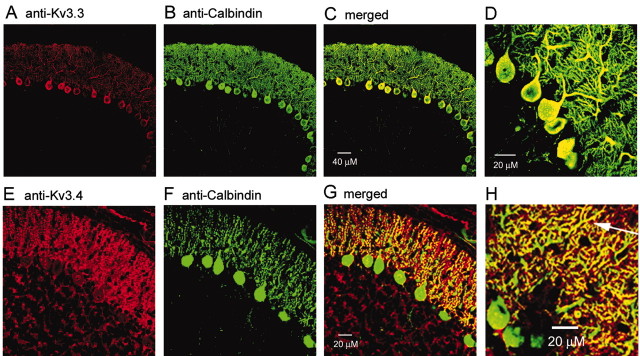
We characterized the properties and functional roles of voltage-dependent potassium channels in the dendrites of Purkinje neurons studied in rat cerebellar slices. Using outside-out patches formed <or=250 microm away from the soma, we found that depolarization-activated potassium channels were present at high density throughout the dendritic tree. Currents required relatively large depolarizations for activation (midpoint, approximately -10 mV), had rapid activation and deactivation kinetics, and inactivated partially (20-70% over 200 msec) with both fast (time constant, 15-20 msec) and slow (300-400 msec) components. Inactivating and noninactivating components were both blocked potently by external tetraethylammonium (half-block by 150 microm) and 4-aminopyridine (half-block by 110 microm). The voltage dependence, kinetics, and pharmacology suggest a predominant contribution by Kv3 family subunits, and immunocytochemical experiments showed staining for both Kv3.3 and Kv3.4 subunits in the dendritic tree. In the proximal dendrite, potassium channels were activated by passively spread sodium spikes recorded at the same position, and experiments using dual recordings showed that the channels serve to actively dampen back-propagation of somatic sodium spikes. In more distal dendrites, potassium currents were activated by voltage waveforms taken from climbing fiber responses, suggesting that they help shape these responses as well. The requirement for large depolarizations allows dendritic Kv3 channels to shape large depolarizing events while not disrupting spatial and temporal summation of smaller excitatory postsynaptic potentials.
Figures






Similar articles
-
Dendritic Kv3.3 potassium channels in cerebellar purkinje cells regulate generation and spatial dynamics of dendritic Ca2+ spikes.J Neurophysiol. 2010 Jun;103(6):3516-25. doi: 10.1152/jn.00982.2009. Epub 2010 Mar 31. J Neurophysiol. 2010. PMID: 20357073 Free PMC article.
-
Kv3 K+ channels enable burst output in rat cerebellar Purkinje cells.Eur J Neurosci. 2004 Aug;20(3):729-39. doi: 10.1111/j.1460-9568.2004.03539.x. Eur J Neurosci. 2004. PMID: 15255983
-
Voltage-dependent potassium currents during fast spikes of rat cerebellar Purkinje neurons: inhibition by BDS-I toxin.J Neurophysiol. 2007 Jan;97(1):563-71. doi: 10.1152/jn.00269.2006. Epub 2006 Oct 25. J Neurophysiol. 2007. PMID: 17065256
-
Kv3 channels: voltage-gated K+ channels designed for high-frequency repetitive firing.Trends Neurosci. 2001 Sep;24(9):517-26. doi: 10.1016/s0166-2236(00)01892-0. Trends Neurosci. 2001. PMID: 11506885 Review.
-
The role of Kv3-type potassium channels in cerebellar physiology and behavior.Cerebellum. 2009 Sep;8(3):323-33. doi: 10.1007/s12311-009-0098-4. Epub 2009 Feb 27. Cerebellum. 2009. PMID: 19247732 Review.
Cited by
-
4-aminopyridine does not enhance flocculus function in tottering, a mouse model of vestibulocerebellar dysfunction and ataxia.PLoS One. 2013;8(2):e57895. doi: 10.1371/journal.pone.0057895. Epub 2013 Feb 25. PLoS One. 2013. PMID: 23451282 Free PMC article.
-
Subunit dependence of Na channel slow inactivation and open channel block in cerebellar neurons.Biophys J. 2007 Mar 15;92(6):1938-51. doi: 10.1529/biophysj.106.093500. Epub 2006 Dec 22. Biophys J. 2007. PMID: 17189307 Free PMC article.
-
Dendritic Kv3.3 potassium channels in cerebellar purkinje cells regulate generation and spatial dynamics of dendritic Ca2+ spikes.J Neurophysiol. 2010 Jun;103(6):3516-25. doi: 10.1152/jn.00982.2009. Epub 2010 Mar 31. J Neurophysiol. 2010. PMID: 20357073 Free PMC article.
-
Action potential processing in a detailed Purkinje cell model reveals a critical role for axonal compartmentalization.Front Cell Neurosci. 2015 Feb 24;9:47. doi: 10.3389/fncel.2015.00047. eCollection 2015. Front Cell Neurosci. 2015. PMID: 25759640 Free PMC article.
-
Interaction of Kv3 potassium channels and resurgent sodium current influences the rate of spontaneous firing of Purkinje neurons.J Neurosci. 2006 Apr 26;26(17):4602-12. doi: 10.1523/JNEUROSCI.5204-05.2006. J Neurosci. 2006. PMID: 16641240 Free PMC article.
References
-
- Callaway JC, Ross WN ( 1997) Spatial distribution of synaptically activated sodium concentration changes in cerebellar Purkinje neurons. J Neurophysiol 77: 145–152. - PubMed
-
- Christie BR, Eliot LS, Ito K, Miyakawa H, Johnston D ( 1995) Different Ca2+ channels in soma and dendrites of hippocampal pyramidal neurons mediate spike-induced Ca2+ influx. J Neurophysiol 73: 2553–2557. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources