

Complex landscape of protein structural dynamics unveiled by nanosecond Laue crystallography
- PMID: 12847289
- PMCID: PMC166376
- DOI: 10.1073/pnas.1430900100
Complex landscape of protein structural dynamics unveiled by nanosecond Laue crystallography
Abstract
Although conformational changes are essential for the function of proteins, little is known about their structural dynamics at atomic level resolution. Myoglobin (Mb) is the paradigm to investigate conformational dynamics because it is a simple globular heme protein displaying a photosensitivity of the iron-ligand bond. Upon laser photodissociation of carboxymyoglobin Mb a nonequilibrium population of protein structures is generated that relaxes over a broad time range extending from picoseconds to milliseconds. This process is associated with migration of the ligand to cavities in the matrix and with a reduction in the geminate rebinding rate by several orders of magnitude. Here we report nanosecond time-resolved Laue diffraction data to 1.55-A resolution on a Mb mutant, which depicts the sequence of structural events associated with this extended relaxation. Motions of the distal E-helix, including the mutated residue Gln-64(E7), and of the CD-turn are found to lag significantly (100-300 ns) behind local rearrangements around the heme such as heme tilting, iron motion out of the heme plane, and swinging of the mutated residue Tyr-29(B10), all of which occur promptly (< or =3 ns). Over the same delayed time range, CO is observed to migrate from a cavity distal to the heme known to bind xenon (called Xe4) to another such cavity proximal to the heme (Xe1). We propose that the extended relaxation of the globin moiety reflects reequilibration among conformational substates known to play an essential role in controlling protein function.
Figures

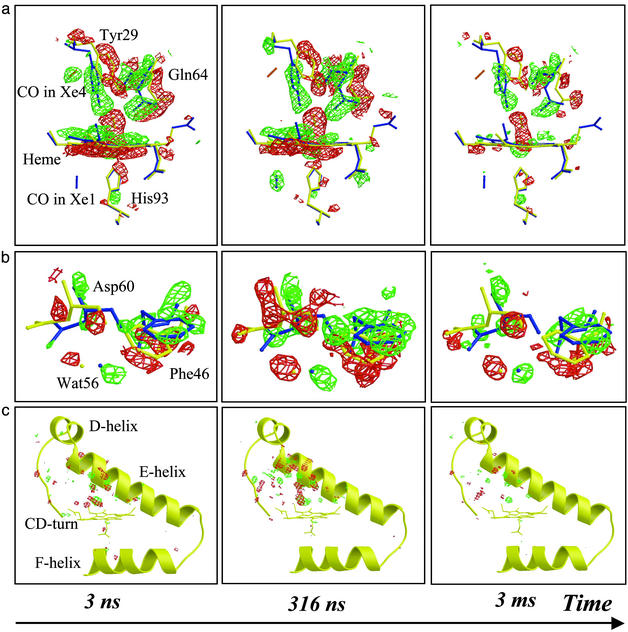
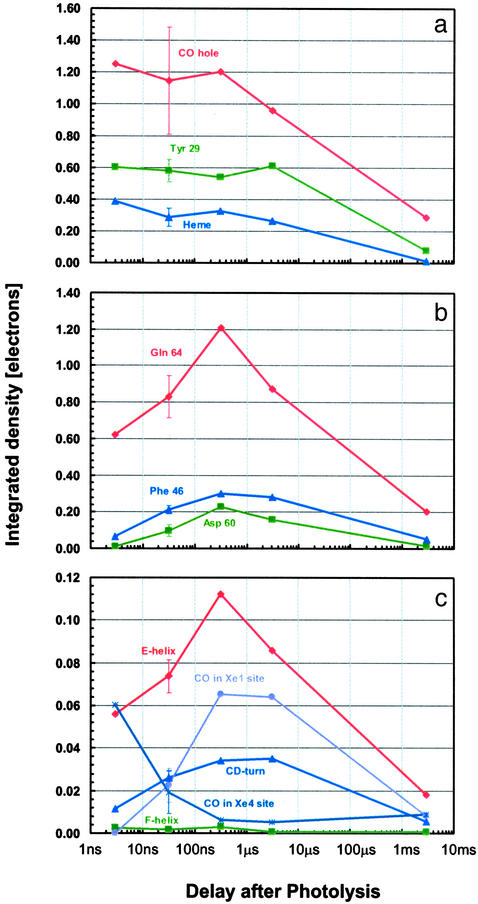
Comment in
-
Myoglobin: the hydrogen atom of biology and a paradigm of complexity.Proc Natl Acad Sci U S A. 2003 Jul 22;100(15):8615-7. doi: 10.1073/pnas.1633688100. Epub 2003 Jul 14. Proc Natl Acad Sci U S A. 2003. PMID: 12861080 Free PMC article. Review. No abstract available.
References
-
- Jackson, T. A., Lim, M. & Anfinrud, P. A. (1994) Chem. Phys. 180, 131–140.
-
- Steinbach, P. J., Ansari, A., Berendsen, J., Braunstein, D., Chu, K., Cowen, B. R., Ehrenstein, D., Frauenfelder H., Johnson, J. B. & Lamb, D. C. (1991) Biochemistry 30, 3988–4001. - PubMed
-
- Ansari, A., Jones, C. M., Henry, E. R., Hofrichter, J. & Eaton, W. A. (1994) Biochemistry 33, 5128–5145. - PubMed
-
- Austin, R. H., Beeson, K. W., Eisenstein, L., Frauenfelder, H. & Gunsalus, I. C. (1975) Biochemistry 14, 5355–5373. - PubMed
-
- Frauenfelder, H., Sligar, S. G. & Wolynes, P. G. (1991) Science 254, 1598–1603. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Miscellaneous
