

Review
doi: 10.1093/emboj/cdg337.
Structure, function and evolution of the signal recognition particle
Affiliations
- PMID: 12853463
- PMCID: PMC165607
- DOI: 10.1093/emboj/cdg337
Item in Clipboard
Review
Structure, function and evolution of the signal recognition particle
EMBO J.
.
Abstract
The signal recognition particle (SRP) is a ribonucleoprotein particle essential for the targeting of signal peptide-bearing proteins to the prokaryotic plasma membrane or the eukaryotic endoplasmic reticulum membrane for secretion or membrane insertion. SRP binds to the signal peptide emerging from the exit site of the ribosome and forms a ribosome nascent chain (RNC)-SRP complex. The RNC-SRP complex then docks in a GTP-dependent manner with a membrane-anchored SRP receptor and the protein is translocated across or integrated into the membrane through a channel called the translocon. Recently considerable progress has been made in understanding the architecture and function of SRP.
Figures
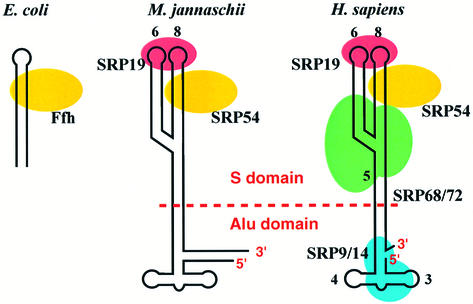
Fig. 1. Schematic representation of SRPs from three kingdoms of life: eubacteria (E.coli, left); archaebacteria (M.jannaschii; middle); and eukarya (Homo sapiens; right). Mammalian SRP can be separated by micrococcal nuclease into two functional domains: the S and Alu domains.
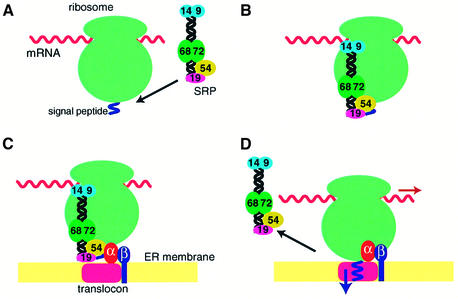
Fig. 2. Functional cycle of the mammalian SRP. (A) SRP binds through the M domain of SRP54 to the signal sequence of membrane and secretory proteins emerging from the exit site of the large ribosomal subunit. (B) The Alu domain promotes transient arrest of the polypeptide chain elongation through an as yet unknown mechanism. The affinity of SRP54 for GTP increases upon docking of SRP with the ribosome. (C) The RNC–SRP complex diffuses to the ER membrane and docks with the SR mainly through the interaction between SRP54 and SRα in the GTP-bound form. SRβ in the GTP-bound form interacts with the RNC complex and induces the transfer of the signal peptide to the translocon. (D) SRP54 and SRα mutually activate their GTPases, and SRP dissociates from the SR upon hydrolysis of GTP, allowing the elongation of the polypeptide to resume.
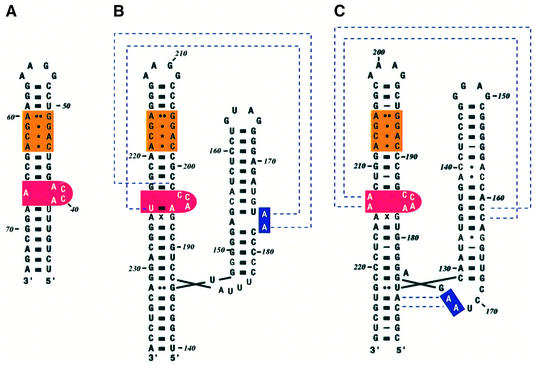
Fig. 3. Secondary structure of SRP RNAs. (A) Escherichia coli, (B) M.jannaschii and (C) H.sapiens. The E.coli 4.5S RNA is 114 nucleotide long and forms an extended hairpin structure. The S domain of M.jannaschii and human 7SL RNA has two branches referred to as helices 6 and 8. Helix 8 bears similarities to domain IV of 4.5S RNA, the binding site for Ffh. The symmetric loop (orange) has the same four non-Watson–Crick base pairs, and the asymmetric loop (red) contains the ACC tri-nucleotide. The dotted lines for M.jannaschii RNA show the interaction between adenines in helix 6 with the backbone in helix 8 found in the SRP19–RNA complex (Hainzl et al., 2002). The dotted lines for human RNA show the A minor motif interactions found in the ternary complex (Kuglstatter et al., 2002).
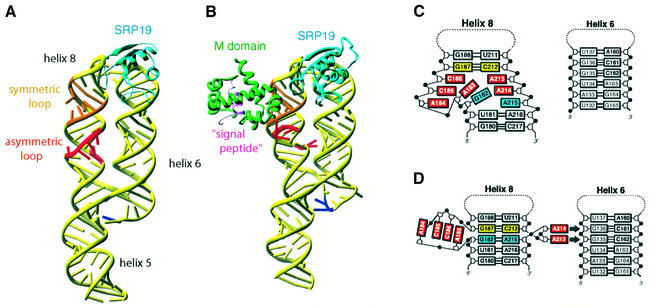
Fig. 4. Conformational changes of the S domain of human 7SL RNA during assembly. (A) The structure of the S domain of human 7SL RNA in complex with M.jannaschii SRP19 (PDB code:1L9A; Oubridge et al., 2002). SRP19 binds to the tetra-loops of helices 6 and 8, clamping them and allowing them to lie side by side. This interaction reduces the flexibility of the asymmetric loop in helix 8 and partially pre-organizes the binding site for SRP54. (B) The binding of the M domain of SRP54 induces a striking conformational change in the structure of the asymmetric loop (PDB code: 1MFQ; Kuglstatter et al., 2002). (C) In the binary complex, the bases of the shorter strand of the asymmetric loop continuously stack, but the longer strand still retains substantial flexibility. (D) In the ternary complex, the asymmetric loop collapses and the G187–C212 base pair (yellow) becomes directly stacked onto the G182–A215 base pair (blue) (Kuglstatter et al., 2002). The bases of A184, C185, C186 and A183 continuously stack, forming a structure reminiscent of the RNA platform seen in the E.coli Ffh–4.5S RNA complex (Batey et al., 2000). A213 and A214 flip out and form A minor motif interactions with the G135–C162 and G136–C161 base pairs of helix 6. This figure was generated using Ribbons (http://sgce.cbse.uab.edu/ribbons/ ). For an animation of the SRP S domain assembly, see http://www.mssm.edu/students/jovinl02/research/srp_assembly_movie.html
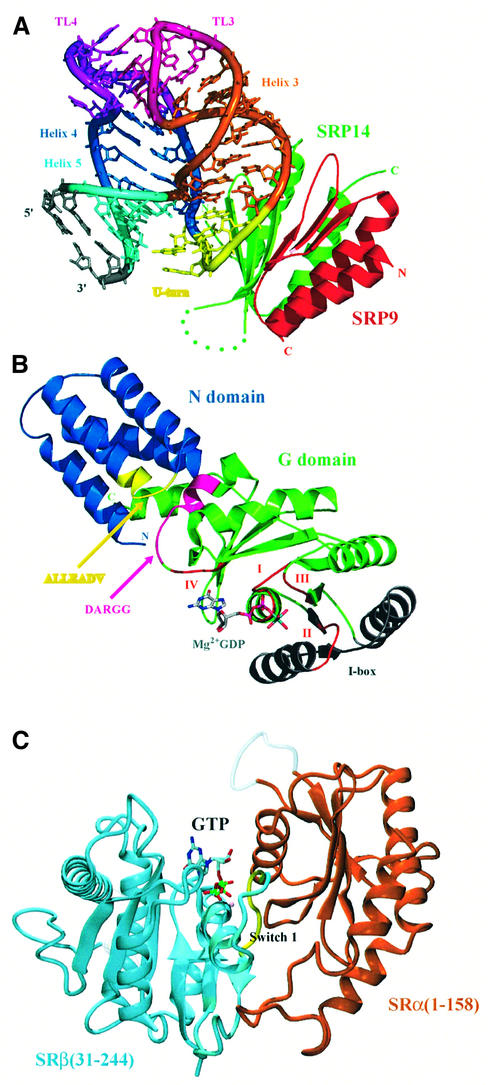
Fig. 5. The crystal structure of the mammalian Alu domain, the NG domain of T.aquaticus Ffh and part of the SR. (A) The crystal structure of SRP9–SRP14 in complex with a small fragment of the Alu domain RNA (PDB code: 1E8O; Weichenrieder et al., 2000). Green, SRP14; red, SRP9; yellow, U-turn; dark blue, helix 4 (H1.2); light blue, helix 5; orange, helix 3. (B) The crystal structure of the Ffh NG domain in the GDP-Mg2+-bound form (PDB code: 1NG1; Freymann et al., 1999). The conserved GTP-binding motifs (I, II, III and IV) are shown in red. The conserved ALLEADV (N domain) and DARGG (G domain) motifs and the I box proposed to be important in coupling the binding of signal peptide, GTP and SRs are highlighted in yellow, purple and grey (Newitt and Bernstein, 1997; Freymann et al., 1999). The actual sequence of the ALLEADV motif in T.aquaticus Ffh is ‘ALMDADV’. (C) The crystal structure of yeast SRβ (blue) in complex with the N-terminal SRβ-interacting (SRX) domain of SRα (brown) (PDB code: 1NRJ; Schwartz and Blobel, 2003). A tightly bound GTP is found at the interface between the two subunits, and the switch I of the GTPase domain of SRβ interacts extensively with SRX (courtesy of Tom Schwartz). (A) and (B) were generated using PyMOL (http://pymol.sourceforge.net/ ).
References
-
- Bacher G., Lütcke,H., Jungnickel,B., Rapoport,T.A. and Dobberstein,B. (1996) Regulation by the ribosome of the GTPase of the signal-recognition particle during protein targeting. Nature, 381, 248–251. - PubMed
-
- Batey R.T., Rambo,R.P., Lucast,L., Rha,B. and Doudna,J.A. (2000) Crystal structure of the ribonucleoprotein core of the signal recognition particle. Science, 287, 1232–1239. - PubMed
-
- Batey R.T., Sagar,M.B. and Doudna,J.A. (2001) Structural and energetic analysis of RNA recognition by a universally conserved protein from the signal recognition particle. J. Mol. Biol., 307, 229–246. - PubMed
-
- Beckmann R., Spahn,C.M.T., Eswar,N., Helmers,J., Penczek,P.A., Sali,A., Frank,J. and Blobel,G. (2001) Architecture of the protein-conducting channel associated with the translating 80S ribosome. Cell, 107, 361–372. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
