

Stimulation of poly(A) polymerase through a direct interaction with the nuclear poly(A) binding protein allosterically regulated by RNA
- PMID: 12853485
- PMCID: PMC165617
- DOI: 10.1093/emboj/cdg347
Stimulation of poly(A) polymerase through a direct interaction with the nuclear poly(A) binding protein allosterically regulated by RNA
Abstract
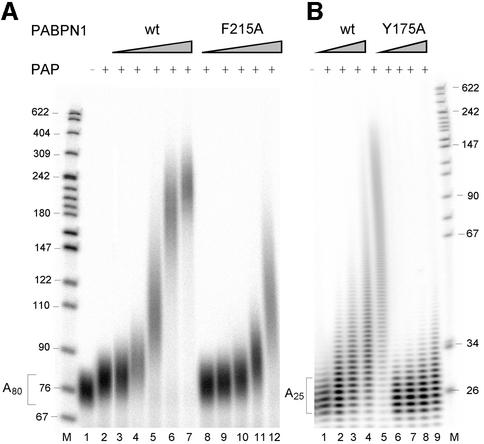
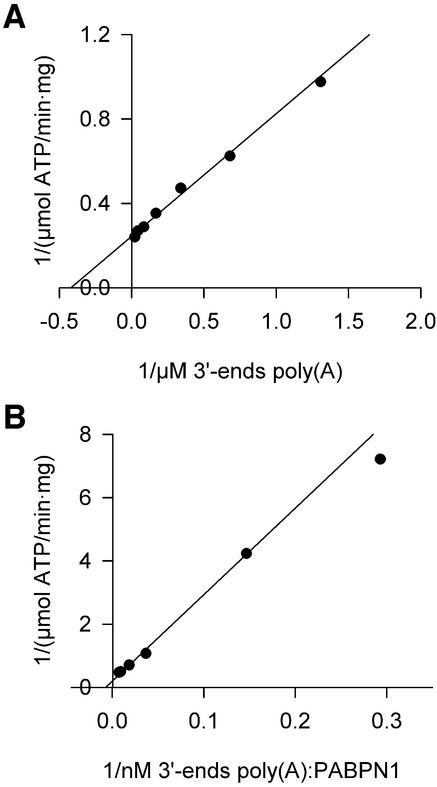
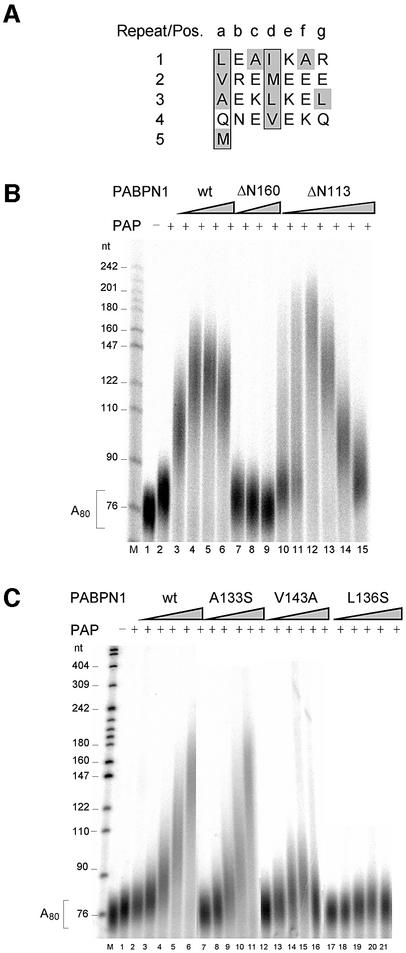
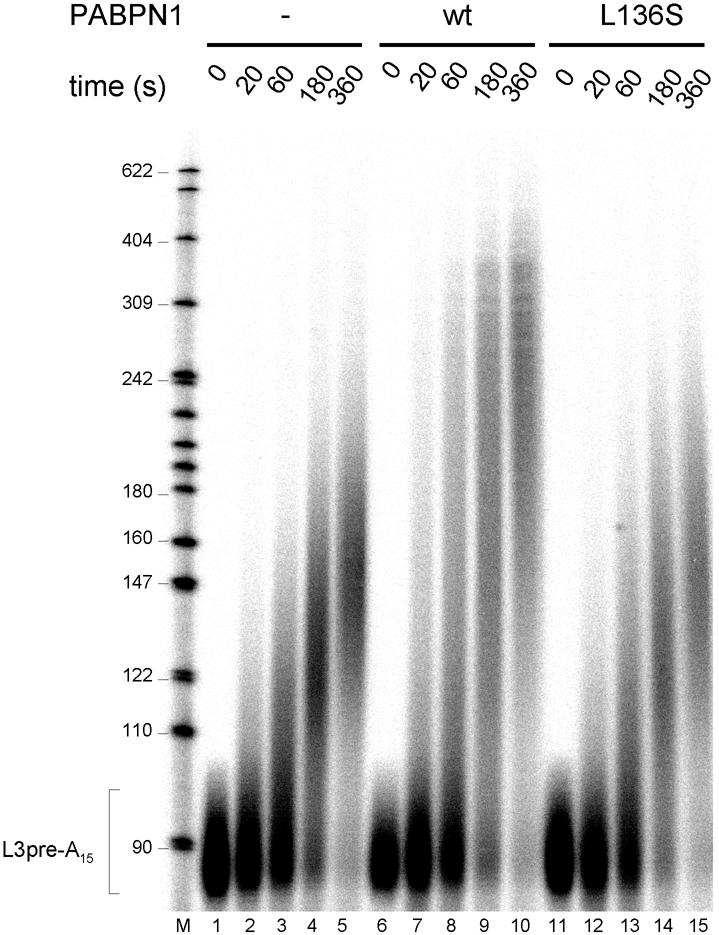
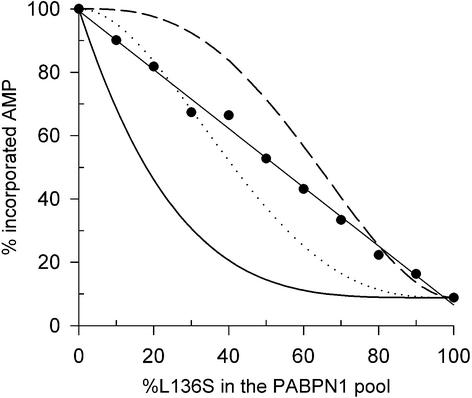
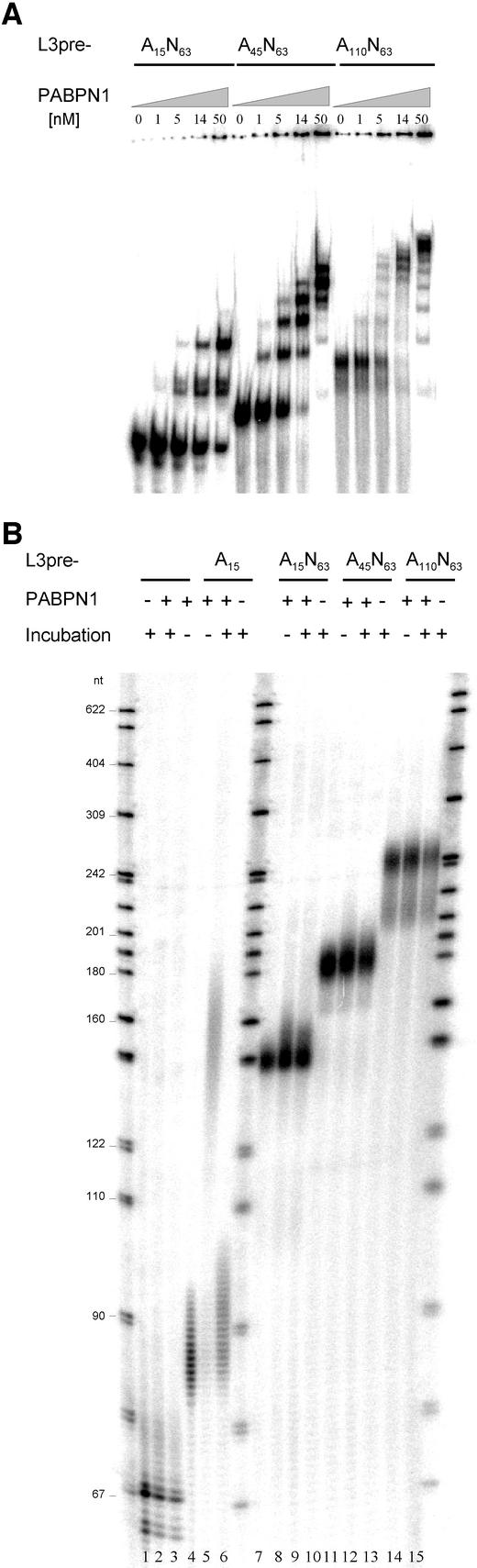
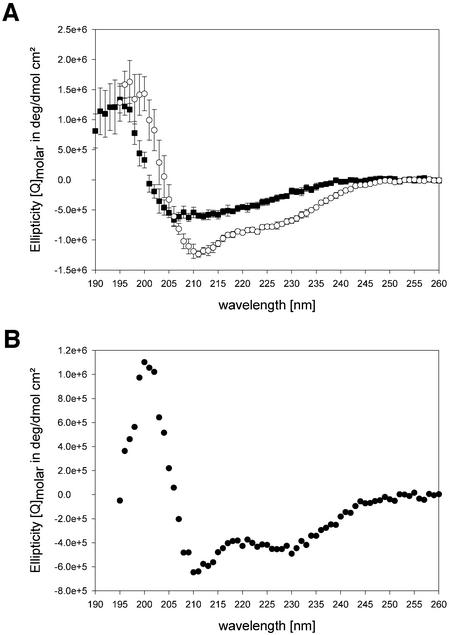
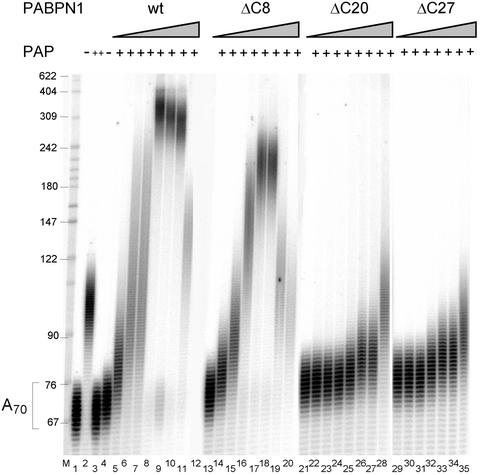
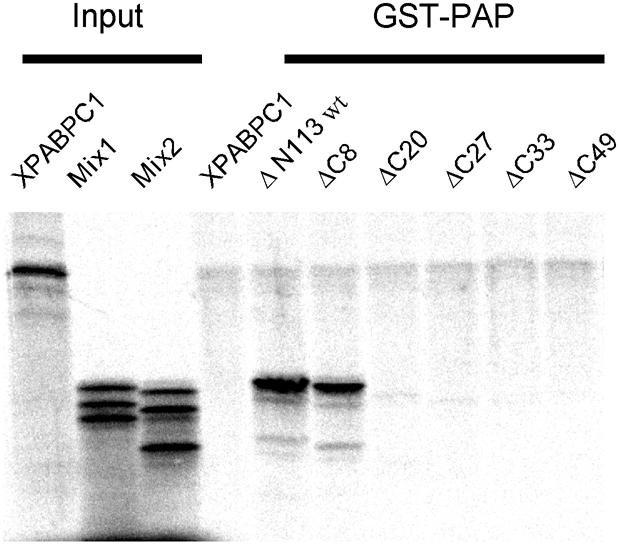
During polyadenylation of mRNA precursors in metazoan cells, poly(A) polymerase is stimulated by the nuclear poly(A) binding protein PABPN1. We report that stimulation depends on binding of PABPN1 to the substrate RNA directly adjacent to poly(A) polymerase and results in an approximately 80-fold increase in the apparent affinity of poly(A) polymerase for RNA without significant effect on catalytic efficiency. PABPN1 associates directly with poly(A) polymerase either upon allosteric activation by oligo(A) or, in the absence of RNA, upon deletion of its N-terminal domain. The N-terminal domain of PABPN1 may function to inhibit undesirable interactions of the protein; the inhibition is relieved upon RNA binding. Tethering of poly(A) polymerase is mediated largely by the C-terminal domain of PABPN1 and is necessary but not sufficient for stimulation of the enzyme; an additional interaction dependent on a coiled-coil structure located within the N-terminal domain of PABPN1 is required for a productive interaction.
Figures
References
-
- Bienroth S., Wahle,E., Suter-Crazzolara,C. and Keller,W. (1991) Purification and characterisation of the cleavage and polyadenylation specificity factor involved in the 3′ processing of messenger RNA precursors. J. Biol. Chem., 266, 19768–19776. - PubMed
-
- Böhm G., Muhr,R. and Jaenicke,R. (1992) Quantitative analysis of protein far UV circular dichroism spectra by neural networks. Protein Eng., 5, 191–195. - PubMed
-
- Bradford M.M. (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem., 72, 248–254. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
