

Early mitotic degradation of the homeoprotein HOXC10 is potentially linked to cell cycle progression
- PMID: 12853486
- PMCID: PMC165610
- DOI: 10.1093/emboj/cdg340
Early mitotic degradation of the homeoprotein HOXC10 is potentially linked to cell cycle progression
Abstract
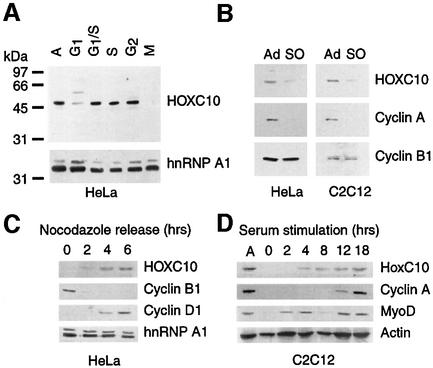
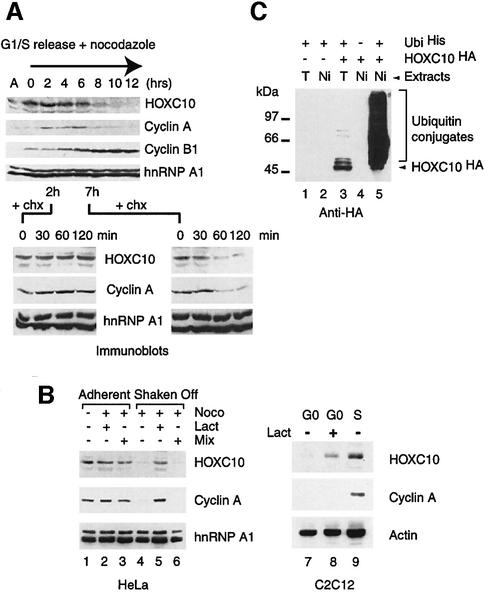
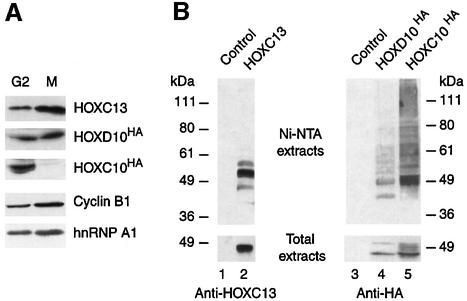
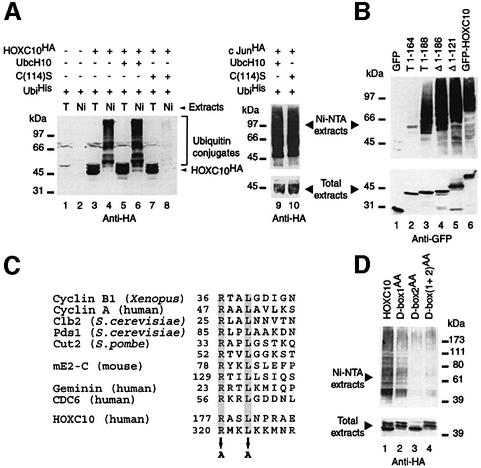
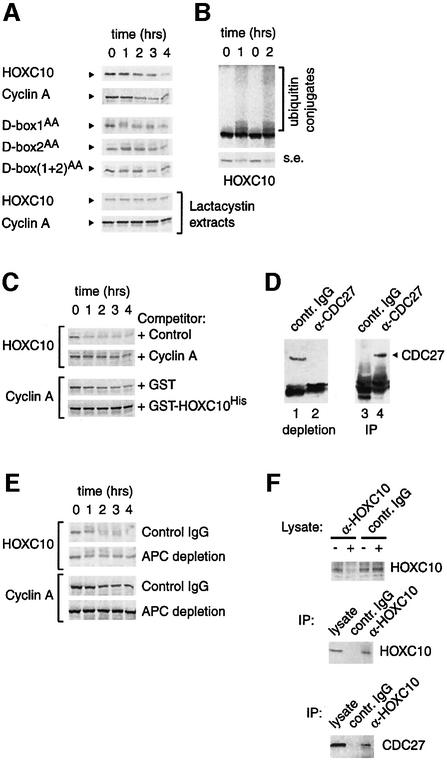
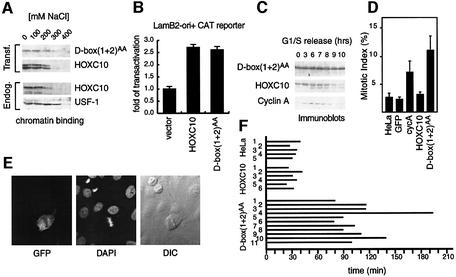
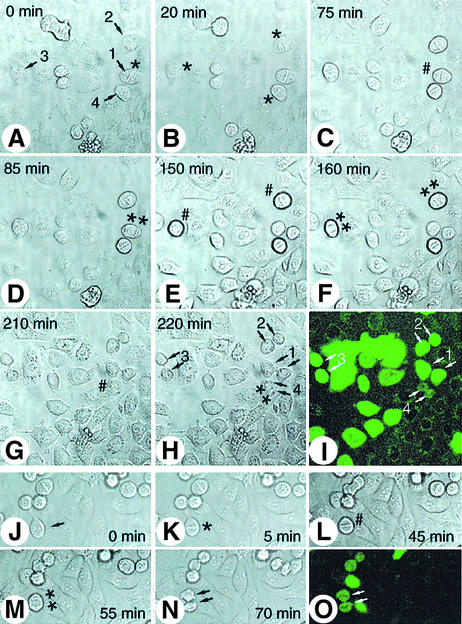
Hox proteins are transcription factors involved in controlling axial patterning, leukaemias and hereditary malformations. Here, we show that HOXC10 oscillates in abundance during the cell cycle, being targeted for degradation early in mitosis by the ubiquitin-dependent proteasome pathway. Among abdominal-B subfamily members, the mitotic proteolysis of HOXC10 appears unique, since the levels of the paralogous HOXD10 and the related homeoprotein HOXC13 are constant throughout the cell cycle. When two destruction box motifs (D-box) are mutated, HOXC10 is stabilized and cells accumulate in metaphase. HOXC10 appears to be a new prometaphase target of the anaphase-promoting complex (APC), since its degradation coincides with cyclin A destruction and is suppressed by expression of a dominant-negative form of UbcH10, an APC-associated ubiquitin-conjugating enzyme. Moreover, HOXC10 co-immunoprecipitates the APC subunit CDC27, and its in vitro degradation is reduced in APC-depleted extracts or by competition with the APC substrate cyclin A. These data imply that HOXC10 is a homeoprotein with the potential to influence mitotic progression, and might provide a link between developmental regulation and cell cycle control.
Figures
Similar articles
-
[Degradation mechanism of cell cycle factors by the proteasome].Nihon Rinsho. 1996 Mar;54(3):861-9. Nihon Rinsho. 1996. PMID: 8904249 Review. Japanese.
-
The Drosophila Bub3 protein is required for the mitotic checkpoint and for normal accumulation of cyclins during G2 and early stages of mitosis.J Cell Sci. 2005 Jan 1;118(Pt 1):187-98. doi: 10.1242/jcs.01602. J Cell Sci. 2005. PMID: 15615783
-
Cell cycle-regulated proteolysis of mitotic target proteins.Mol Biol Cell. 1999 Nov;10(11):3927-41. doi: 10.1091/mbc.10.11.3927. Mol Biol Cell. 1999. PMID: 10564281 Free PMC article.
-
RNA interference of valosin-containing protein (VCP/p97) reveals multiple cellular roles linked to ubiquitin/proteasome-dependent proteolysis.J Cell Sci. 2004 Jan 15;117(Pt 2):281-92. doi: 10.1242/jcs.00841. Epub 2003 Dec 2. J Cell Sci. 2004. PMID: 14657277
-
[Regulation of cell cycle progression by proteolysis].Tanpakushitsu Kakusan Koso. 1996 Sep;41(12 Suppl):1820-5. Tanpakushitsu Kakusan Koso. 1996. PMID: 8890642 Review. Japanese. No abstract available.
Cited by
-
The homeodomain transcription factor Hoxa2 interacts with and promotes the proteasomal degradation of the E3 ubiquitin protein ligase RCHY1.PLoS One. 2013 Nov 7;8(11):e80387. doi: 10.1371/journal.pone.0080387. eCollection 2013. PLoS One. 2013. PMID: 24244684 Free PMC article.
-
Molecular mechanisms of APC/C release from spindle assembly checkpoint inhibition by APC/C SUMOylation.Cell Rep. 2021 Mar 30;34(13):108929. doi: 10.1016/j.celrep.2021.108929. Cell Rep. 2021. PMID: 33789095 Free PMC article.
-
Transcriptome analysis revealed unique genes as targets for the anti-inflammatory action of activated protein C in human macrophages.PLoS One. 2010 Oct 15;5(10):e15352. doi: 10.1371/journal.pone.0015352. PLoS One. 2010. PMID: 20976180 Free PMC article.
-
A Numb-Mdm2 fuzzy complex reveals an isoform-specific involvement of Numb in breast cancer.J Cell Biol. 2018 Feb 5;217(2):745-762. doi: 10.1083/jcb.201709092. Epub 2017 Dec 21. J Cell Biol. 2018. PMID: 29269425 Free PMC article.
-
APC/C(Cdc20) targets E2F1 for degradation in prometaphase.Cell Cycle. 2010 Oct 1;9(19):3956-64. doi: 10.4161/cc.9.19.13162. Epub 2010 Oct 26. Cell Cycle. 2010. PMID: 20948288 Free PMC article.
References
-
- Abate-Shen C. (2002) Deregulated homeobox gene expression in cancer: cause or consequence? Nat. Rev. Cancer, 2, 777–785. - PubMed
-
- Abdurashidova G., Deganuto,M., Klima,R., Riva,S., Biamonti,G., Giacca,M. and Falaschi,A. (2000) Start sites of bidirectional DNA synthesis at the human lamin B2 origin. Science, 287, 2023–2026. - PubMed
-
- Boncinelli E. (1997) Homeobox genes and disease. Curr. Opin. Genet. Dev., 7, 331–337. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
