

The ErbB2/ErbB3 heterodimer functions as an oncogenic unit: ErbB2 requires ErbB3 to drive breast tumor cell proliferation
- PMID: 12853564
- PMCID: PMC166416
- DOI: 10.1073/pnas.1537685100
The ErbB2/ErbB3 heterodimer functions as an oncogenic unit: ErbB2 requires ErbB3 to drive breast tumor cell proliferation
Abstract
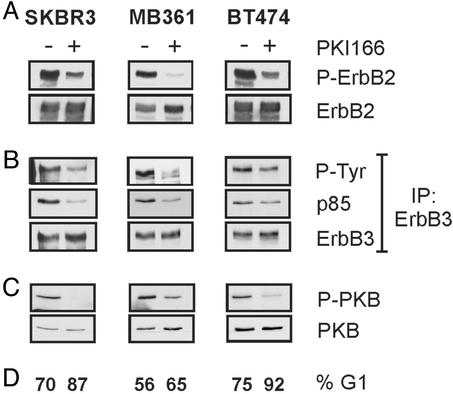
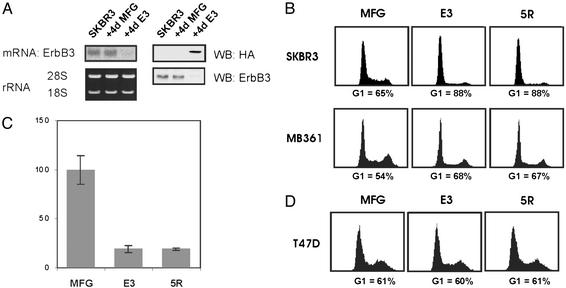
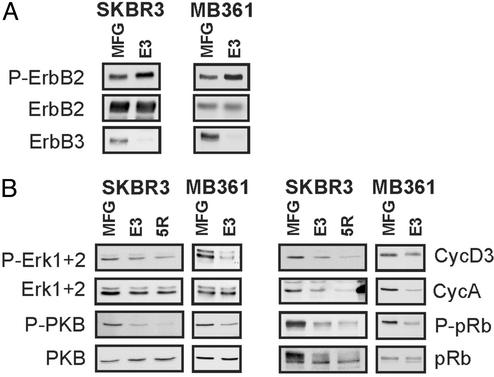
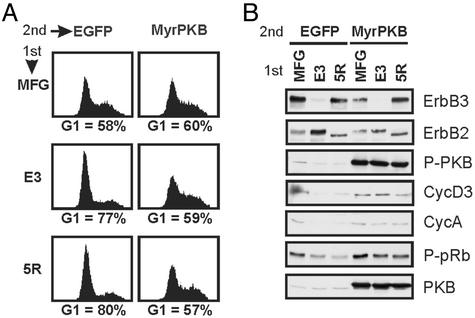
ErbB2 is a receptor tyrosine kinase whose activity in normal cells depends on dimerization with another ligand-binding ErbB receptor. In contrast, amplification of c-erbB2 in tumors results in dramatic overexpression and constitutive activation of the receptor. Breast cancer cells overexpressing ErbB2 depend on its activity for proliferation, because treatment of these cells with ErbB2-specific antagonistic antibodies or kinase inhibitors blocks tumor cells in the G1 phase of the cell cycle. Intriguingly, loss of ErbB2 signaling is accompanied by a decrease in the phosphotyrosine content of ErbB3. On the basis of these results, it has been proposed that ErbB3 might be a partner for ErbB2 in promoting cellular transformation. To test this hypothesis and directly examine the role of the "kinase dead" ErbB3, we specifically ablated its expression with a designer transcription factor (E3). By infection of ErbB2-overexpressing breast cancer cells with a retrovirus expressing E3, we show that ErbB3 is an essential partner in the transformation process. Loss of functional ErbB2 or ErbB3 has similar effects on cell proliferation and cell cycle regulators. Furthermore, expression of constitutively active protein kinase B rescues the proliferative block induced as a consequence of loss of ErbB2 or ErbB3 signaling. These results demonstrate that ErbB2 overexpression and activity alone are insufficient to promote breast tumor cell division. Furthermore, we identify ErbB3's role, which is to couple active ErbB2 to the phosphatidylinositol 3-kinase/protein kinase B pathway. Thus, the ErbB2/ErbB3 dimer functions as an oncogenic unit to drive breast tumor cell proliferation.
Figures

References
-
- Yarden, Y. & Sliwkowski, M. X. (2001) Nat. Rev. Mol. Cell Biol. 2, 127–137. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
