

Human inhibitory receptors Ig-like transcript 2 (ILT2) and ILT4 compete with CD8 for MHC class I binding and bind preferentially to HLA-G
- PMID: 12853576
- PMCID: PMC166403
- DOI: 10.1073/pnas.1431057100
Human inhibitory receptors Ig-like transcript 2 (ILT2) and ILT4 compete with CD8 for MHC class I binding and bind preferentially to HLA-G
Abstract
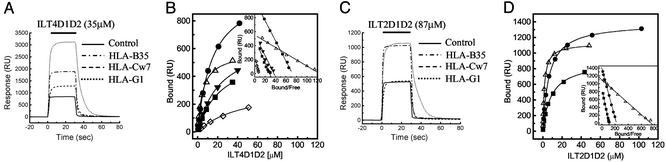
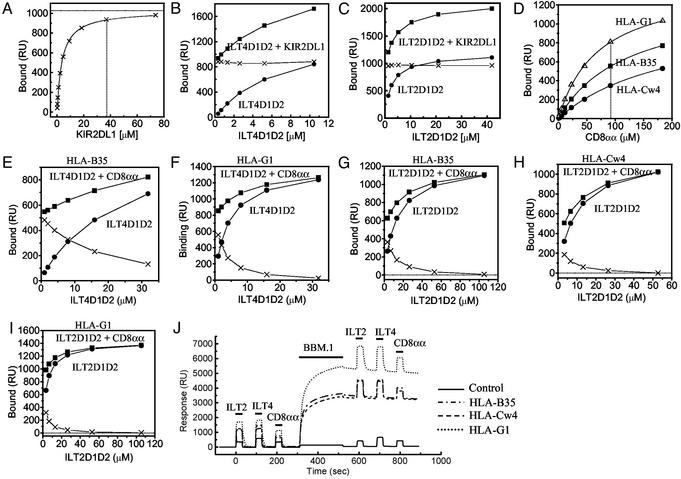
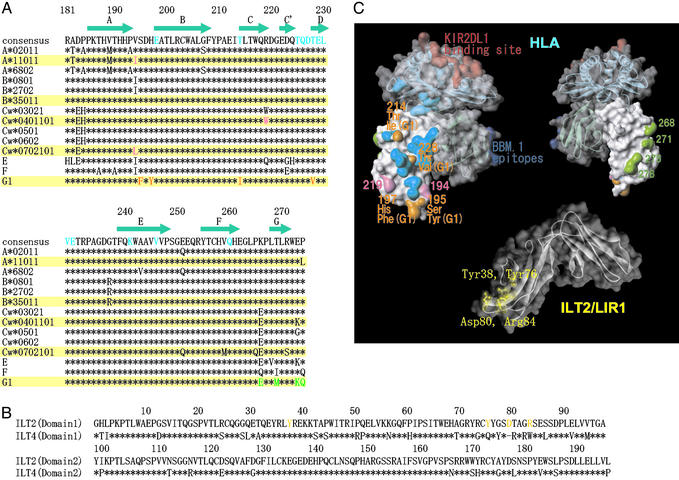
Ig-like transcript 4 (ILT4) (also known as leukocyte Ig-like receptor 2, CD85d, and LILRB2) is a cell surface receptor expressed mainly on myelomonocytic cells, whereas ILT2 (also known as leukocyte Ig-like receptor 1, CD85j, and LILRB1) is expressed on a wider range of immune cells including subsets of natural killer and T cells. Both ILTs contain immunoreceptor tyrosine-based inhibitory receptor motifs in their cytoplasmic tails that inhibit cellular responses by recruiting phosphatases such as SHP-1 (Src homology 2 domain containing tyrosine phosphatase 1). Although these ILTs have been shown to recognize a broad range of classical and nonclassical human MHC class I molecules (MHCIs), their precise binding properties remain controversial. We have used surface plasmon resonance to analyze the interaction of soluble forms of ILT4 and ILT2 with several MHCIs. Although the range of affinities measured was quite broad (Kd = 2-45 microM), some interesting differences were observed. ILT2 generally bound with a 2- to 3-fold higher affinity than ILT4 to the same MHCI. Furthermore, ILT2 and ILT4 bound to HLA-G with a 3- to 4-fold higher affinity than to classical MHCIs, suggesting that ILT/HLA-G recognition may play a dominant role in the regulation of natural killer, T, and myelomonocytic cell activation. Finally, we show that ILT2 and ILT4 effectively compete with CD8 for MHCI binding, raising the possibility that ILT2 modulates CD8+ T cell activation by blocking the CD8 binding as well as by recruiting inhibitory molecules through its immunoreceptor tyrosine-based inhibitory receptor motif.
Figures
References
-
- Martin, A. M., Kulski, J. K., Witt, C., Pontarotti, P. & Christiansen, F. T. (2002) Trends Immunol. 23, 81–88. - PubMed
-
- Samaridis, J. & Collona, M. (1997) Eur. J. Immunol. 27, 660–665. - PubMed
-
- Cosman, D., Fanger, N., Borges, L., Kubin, M., Chin, W., Peterson, L. & Hsu, M.-L. (1997) Immunity 7, 273–282. - PubMed
-
- Borges, L., Hsu, M.-L., Fanger, N., Kubin, M. & Cosman, D. (1997) J. Immunol. 159, 5192–5196. - PubMed
-
- Wagtmann, N., Rojo, S., Eichler, E., Mohrenweiser, H. & Long, O. E. (1997) Curr. Biol. 7, 615–618. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
