

X-ray crystal structures of the WT and a hyper-accurate ribosome from Escherichia coli
- PMID: 12853578
- PMCID: PMC166372
- DOI: 10.1073/pnas.1133380100
X-ray crystal structures of the WT and a hyper-accurate ribosome from Escherichia coli
Abstract
Protein biosynthesis on the ribosome requires accurate reading of the genetic code in mRNA. Two conformational rearrangements in the small ribosomal subunit, a closing of the head and body around the incoming tRNA and an RNA helical switch near the mRNA decoding site, have been proposed to select for complementary base-pairing between mRNA codons and tRNA anticodons. We determined x-ray crystal structures of the WT and a hyper-accurate variant of the Escherichia coli ribosome at resolutions of 10 and 9 A, respectively, revealing that formation of the intact 70S ribosome from its two subunits closes the conformation of the head of the small subunit independent of mRNA decoding. Moreover, no change in the conformation of the switch helix is observed in two steps of tRNA discrimination. These 70S ribosome structures indicate that mRNA decoding is coupled primarily to movement of the small subunit body, consistent with previous proposals, whereas closing of the head and the helical switch may function in other steps of protein synthesis.
Figures
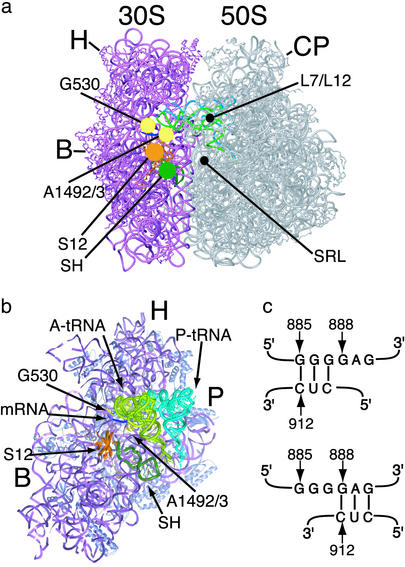
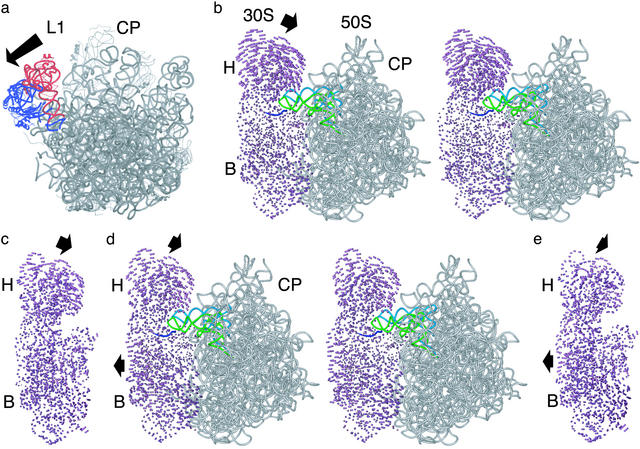
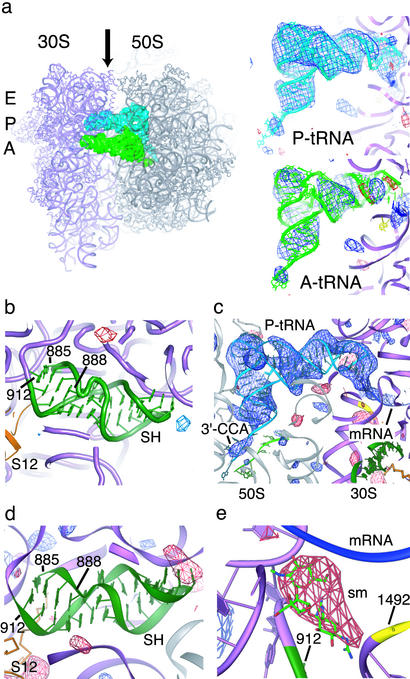
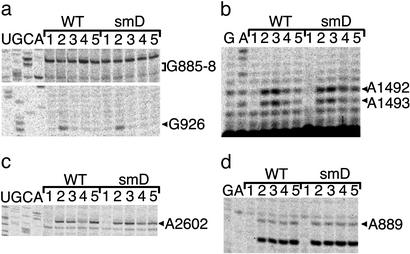
References
-
- Rodnina, M. V. & Wintermeyer, W. (2001) Annu. Rev. Biochem. 70, 415–435. - PubMed
-
- Ogle, J. M., Carter, A. P. & Ramakrishnan, V. (2003) Trends Biochem. Sci. 28, 259–266. - PubMed
-
- Moazed, D. & Noller, H. F. (1986) Cell 47, 985–994. - PubMed
-
- Moazed, D. & Noller, H. F. (1987) Nature 327, 389–394. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
