

Mycobacterium tuberculosis RecA intein, a LAGLIDADG homing endonuclease, displays Mn(2+) and DNA-dependent ATPase activity
- PMID: 12853636
- PMCID: PMC167636
- DOI: 10.1093/nar/gkg475
Mycobacterium tuberculosis RecA intein, a LAGLIDADG homing endonuclease, displays Mn(2+) and DNA-dependent ATPase activity
Abstract
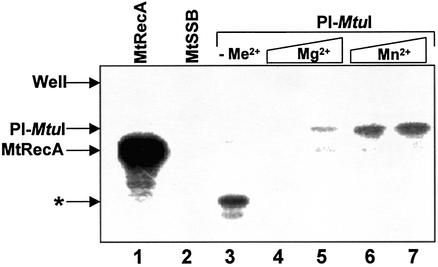
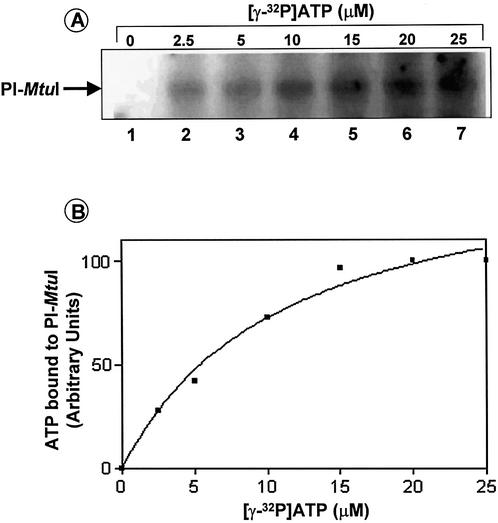
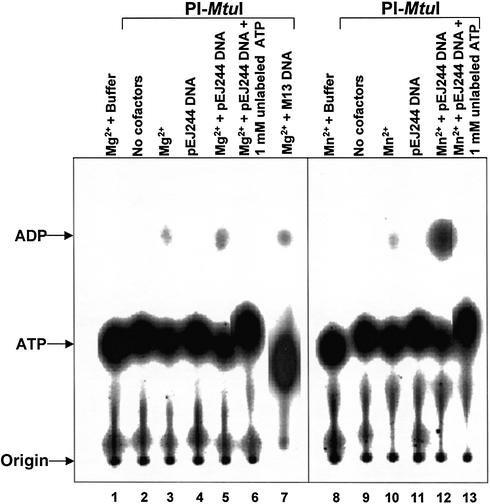
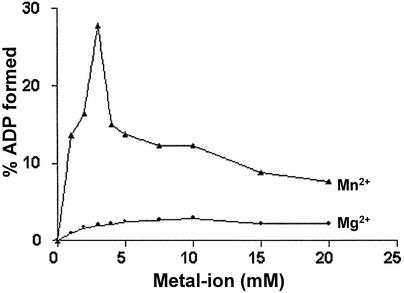
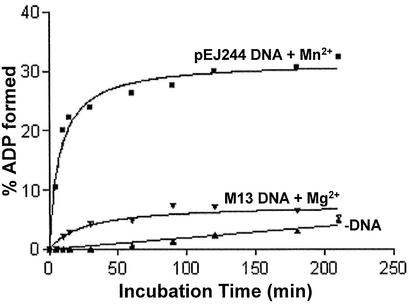
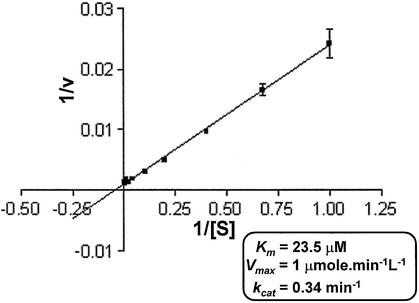
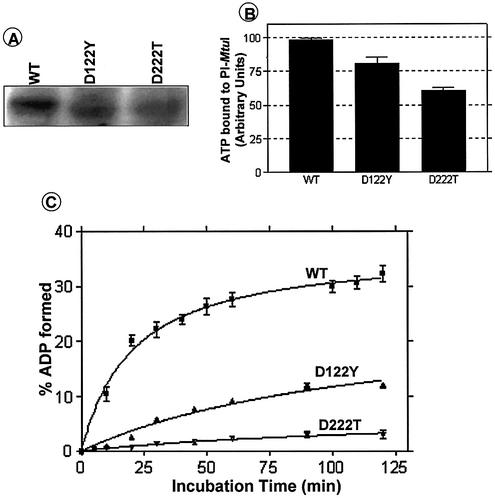
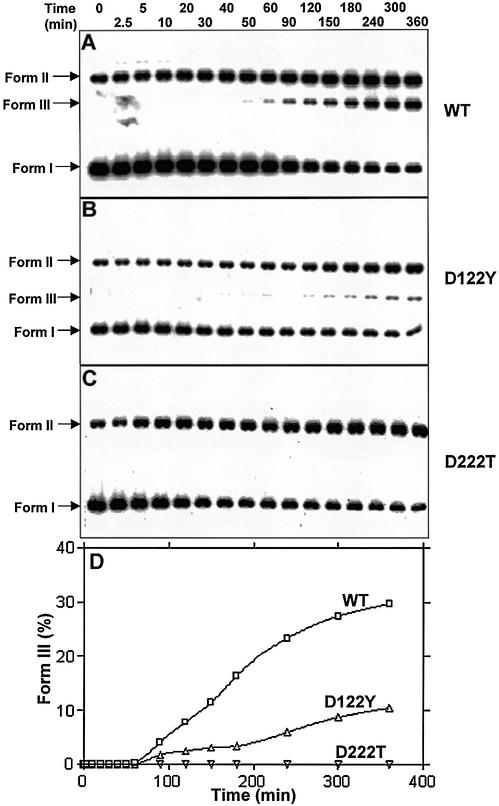
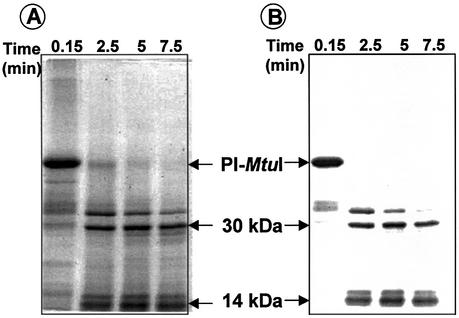
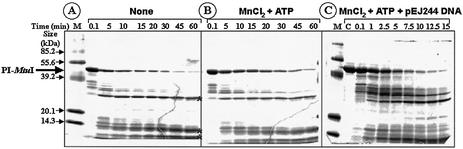
Mycobacterium tuberculosis RecA intein (PI-MtuI), a LAGLIDADG homing endonuclease, displays dual target specificity in response to alternative cofactors. While both ATP and Mn(2+) were required for optimal cleavage of an inteinless recA allele (hereafter referred to as cognate DNA), Mg(2+) alone was sufficient for cleavage of ectopic DNA sites. In this study, we have explored the ability of PI-MtuI to catalyze ATP hydrolysis in the presence of alternative metal ion cofactors and DNA substrates. Our results indicate that PI-MtuI displays maximum ATPase activity in the presence of cognate but not ectopic DNA. Kinetic analysis revealed that Mn(2+) was able to stimulate PI-MtuI catalyzed ATP hydrolysis, whereas Mg(2+) failed to do so. Using UV crosslinking, limited proteolysis and amino acid sequence analysis, we show that (32)P-labeled ATP was bound to a 14 kDa peptide containing the putative Walker A motif. Furthermore, the limited proteolysis approach disclosed that cognate DNA was able to induce structural changes in PI-MtuI. Mutation of the presumptive metal ion-binding ligands (Asp122 and Asp222) in the LAGLIDADG motifs of PI-MtuI impaired its affinity for ATP, thus resulting in a reduction in or loss of its endonuclease activity. Together, these results suggest that PI-MtuI is a (cognate) DNA- and Mn(2+)-dependent ATPase, unique from the LAGLIDADG family of homing endonucleases, and implies a possible role for ATP hydrolysis in the recognition and/or cleavage of homing site DNA sequence.
Figures
Similar articles
-
Characterization of Mycobacterium leprae RecA intein, a LAGLIDADG homing endonuclease, reveals a unique mode of DNA binding, helical distortion, and cleavage compared with a canonical LAGLIDADG homing endonuclease.J Biol Chem. 2009 Sep 18;284(38):25912-28. doi: 10.1074/jbc.M109.042861. Epub 2009 Jul 15. J Biol Chem. 2009. PMID: 19605345 Free PMC article.
-
Mycobacterium tuberculosis RecA intein possesses a novel ATP-dependent site-specific double-stranded DNA endonuclease activity.J Biol Chem. 2002 May 3;277(18):16257-64. doi: 10.1074/jbc.M112365200. Epub 2002 Feb 15. J Biol Chem. 2002. PMID: 11850426
-
The RecA intein of Mycobacterium tuberculosis promotes cleavage of ectopic DNA sites. Implications for the dispersal of inteins in natural populations.J Biol Chem. 2002 Oct 25;277(43):40352-61. doi: 10.1074/jbc.M205697200. Epub 2002 Aug 6. J Biol Chem. 2002. PMID: 12167644
-
Structure and mechanism of Escherichia coli RecA ATPase.Mol Microbiol. 2005 Oct;58(2):358-66. doi: 10.1111/j.1365-2958.2005.04876.x. Mol Microbiol. 2005. PMID: 16194225 Review.
-
How strand exchange protein function benefits from ATP hydrolysis.Curr Opin Genet Dev. 2021 Dec;71:120-128. doi: 10.1016/j.gde.2021.06.016. Epub 2021 Jul 31. Curr Opin Genet Dev. 2021. PMID: 34343922 Free PMC article. Review.
Cited by
-
Conditional protein splicing of the Mycobacterium tuberculosis RecA intein in its native host.Sci Rep. 2024 Sep 5;14(1):20664. doi: 10.1038/s41598-024-71248-y. Sci Rep. 2024. PMID: 39237639 Free PMC article.
-
Conditional protein splicing of the Mycobacterium tuberculosis RecA intein in its native host.bioRxiv [Preprint]. 2024 Apr 15:2024.04.15.589443. doi: 10.1101/2024.04.15.589443. bioRxiv. 2024. Update in: Sci Rep. 2024 Sep 5;14(1):20664. doi: 10.1038/s41598-024-71248-y. PMID: 38659745 Free PMC article. Updated. Preprint.
-
WhiA transcription factor provides feedback loop between translation and energy production in a genome-reduced bacterium.Front Microbiol. 2024 Dec 23;15:1504418. doi: 10.3389/fmicb.2024.1504418. eCollection 2024. Front Microbiol. 2024. PMID: 39764455 Free PMC article.
-
Intein Clustering Suggests Functional Importance in Different Domains of Life.Mol Biol Evol. 2016 Mar;33(3):783-99. doi: 10.1093/molbev/msv271. Epub 2015 Nov 25. Mol Biol Evol. 2016. PMID: 26609079 Free PMC article.
-
Characterization of Mycobacterium leprae RecA intein, a LAGLIDADG homing endonuclease, reveals a unique mode of DNA binding, helical distortion, and cleavage compared with a canonical LAGLIDADG homing endonuclease.J Biol Chem. 2009 Sep 18;284(38):25912-28. doi: 10.1074/jbc.M109.042861. Epub 2009 Jul 15. J Biol Chem. 2009. PMID: 19605345 Free PMC article.
References
-
- Liu X.Q. (2000) Protein-splicing intein: genetic mobility, origin and evolution. Annu. Rev. Genet., 34, 61–76. - PubMed
-
- Noren C.J., Wang,J. and Perler,F.B. (2000) Dissecting the chemistry of protein splicing and its applications. Angew. Chem. Int. Ed. Engl., 39, 450–466. - PubMed
-
- Gimble F.S. (2000) Invasion of a multitude of genetic niches by mobile endonuclease genes. FEMS Microbiol. Lett., 185, 99–107. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
