

Molecular modeling of the chromatosome particle
- PMID: 12853645
- PMCID: PMC167642
- DOI: 10.1093/nar/gkg481
Molecular modeling of the chromatosome particle
Abstract
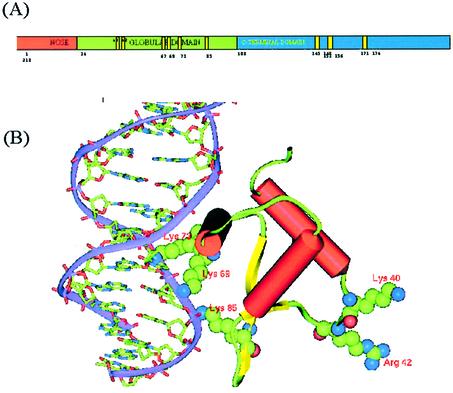
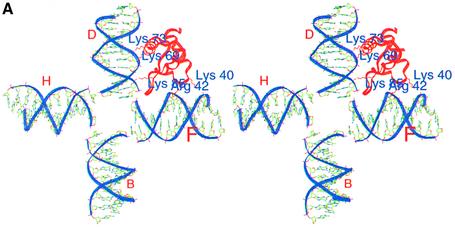
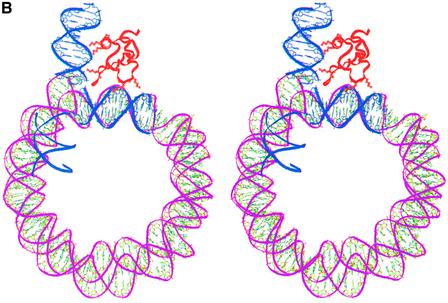
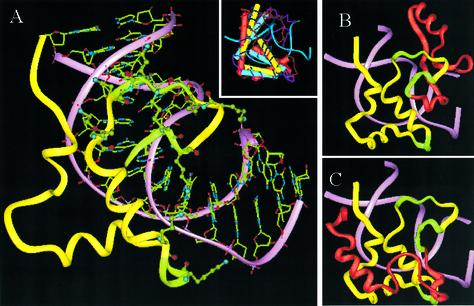
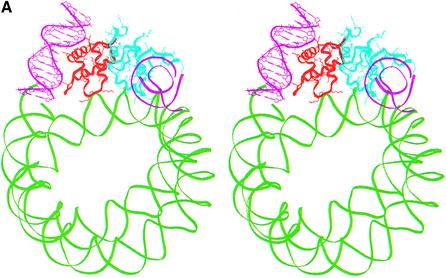
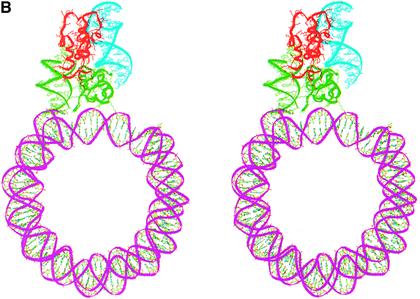
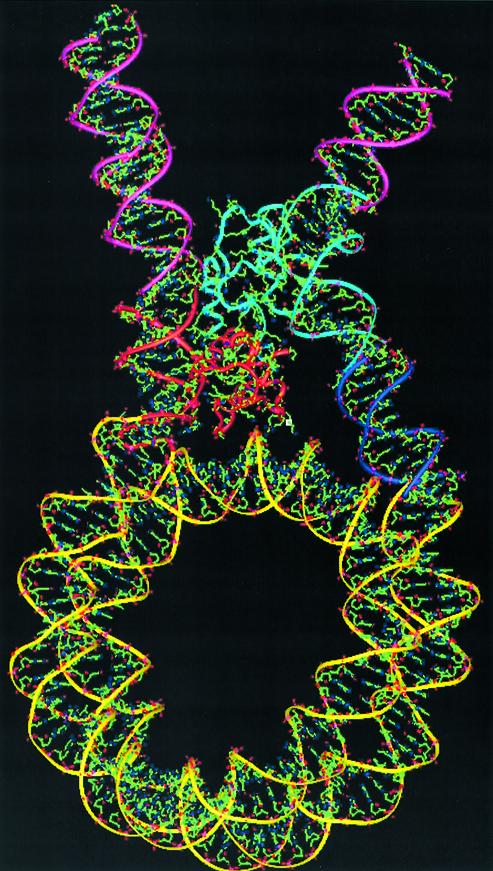
In an effort to understand the role of the linker histone in chromatin folding, its structure and location in the nucleosome has been studied by molecular modeling methods. The structure of the globular domain of the rat histone H1d, a highly conserved part of the linker histone, built by homology modeling methods, revealed a three-helical bundle fold that could be described as a helix-turn-helix variant with its characteristic properties of binding to DNA at the major groove. Using the information of its preferential binding to four-way Holliday junction (HJ) DNA, a model of the domain complexed to HJ was built, which was subsequently used to position the globular domain onto the nucleosome. The model revealed that the primary binding site of the domain interacts with the extra 20 bp of DNA of the entering duplex at the major groove while the secondary binding site interacts with the minor groove of the central gyre of the DNA superhelix of the nucleosomal core. The positioning of the globular domain served as an anchor to locate the C-terminal domain onto the nucleosome to obtain the structure of the chromatosome particle. The resulting structure had a stem-like appearance, resembling that observed by electron microscopic studies. The C-terminal domain which adopts a high mobility group (HMG)-box-like fold, has the ability to bend DNA, causing DNA condensation or compaction. It was observed that the three S/TPKK motifs in the C-terminal domain interact with the exiting duplex, thus defining the path of linker DNA in the chromatin fiber. This study has provided an insight into the probable individual roles of globular and the C-terminal domains of histone H1 in chromatin organization.
Figures
Similar articles
-
Two DNA-binding sites on the globular domain of histone H5 are required for binding to both bulk and 5 S reconstituted nucleosomes.J Mol Biol. 2000 Nov 17;304(1):21-33. doi: 10.1006/jmbi.2000.4205. J Mol Biol. 2000. PMID: 11071807
-
HMGN1 and 2 remodel core and linker histone tail domains within chromatin.Nucleic Acids Res. 2017 Sep 29;45(17):9917-9930. doi: 10.1093/nar/gkx579. Nucleic Acids Res. 2017. PMID: 28973435 Free PMC article.
-
Mapping the interaction surface of linker histone H1(0) with the nucleosome of native chromatin in vivo.Nat Struct Mol Biol. 2006 Mar;13(3):250-5. doi: 10.1038/nsmb1050. Epub 2006 Feb 5. Nat Struct Mol Biol. 2006. PMID: 16462749 Free PMC article.
-
Emerging roles of linker histones in regulating chromatin structure and function.Nat Rev Mol Cell Biol. 2018 Mar;19(3):192-206. doi: 10.1038/nrm.2017.94. Epub 2017 Oct 11. Nat Rev Mol Cell Biol. 2018. PMID: 29018282 Free PMC article. Review.
-
Chromatin structures condensed by linker histones.Essays Biochem. 2019 Apr 23;63(1):75-87. doi: 10.1042/EBC20180056. Print 2019 Apr 23. Essays Biochem. 2019. PMID: 31015384 Review.
Cited by
-
Conformational selection and dynamic adaptation upon linker histone binding to the nucleosome.Nucleic Acids Res. 2016 Aug 19;44(14):6599-613. doi: 10.1093/nar/gkw514. Epub 2016 Jun 7. Nucleic Acids Res. 2016. PMID: 27270081 Free PMC article.
-
Nucleosome assembly protein-1 is a linker histone chaperone in Xenopus eggs.Proc Natl Acad Sci U S A. 2005 Jun 7;102(23):8210-5. doi: 10.1073/pnas.0500822102. Epub 2005 May 31. Proc Natl Acad Sci U S A. 2005. PMID: 15928086 Free PMC article.
-
Complex of linker histone H5 with the nucleosome and its implications for chromatin packing.Proc Natl Acad Sci U S A. 2006 May 30;103(22):8384-9. doi: 10.1073/pnas.0508951103. Epub 2006 May 22. Proc Natl Acad Sci U S A. 2006. PMID: 16717183 Free PMC article.
-
Dependence of Chromatosome Structure on Linker Histone Sequence and Posttranslational Modification.Biophys J. 2018 May 22;114(10):2363-2375. doi: 10.1016/j.bpj.2018.04.034. Epub 2018 May 11. Biophys J. 2018. PMID: 29759374 Free PMC article.
-
Nucleosome structure and function.Chem Rev. 2015 Mar 25;115(6):2255-73. doi: 10.1021/cr500373h. Epub 2014 Dec 12. Chem Rev. 2015. PMID: 25495456 Free PMC article. Review. No abstract available.
References
-
- van Holde K.E. (1988) Chromatin. Spring Verlag, New York.
-
- Wolffe A.P. (1998) Chromatin Structure and Function. Academic Press, London.
-
- Felsenfeld G. (1992) Chromatin as an essential part of the transcriptional mechanism. Nature, 355, 219–223. - PubMed
-
- Fedor M.J. (1992) Chromatin structure and gene expression. Curr. Opin. Cell Biol., 4, 436–443. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
