

Inhibition of human cytomegalovirus DNA polymerase by C-terminal peptides from the UL54 subunit
- PMID: 12857903
- PMCID: PMC165265
- DOI: 10.1128/jvi.77.15.8336-8344.2003
Inhibition of human cytomegalovirus DNA polymerase by C-terminal peptides from the UL54 subunit
Abstract
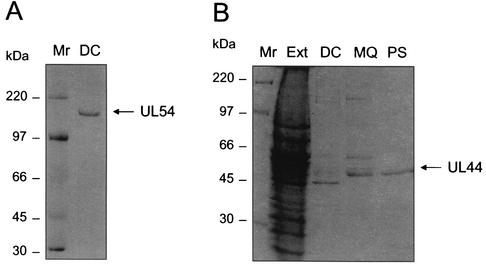
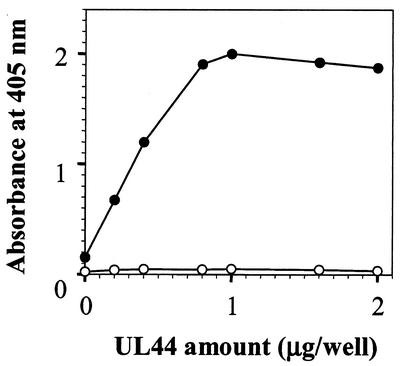
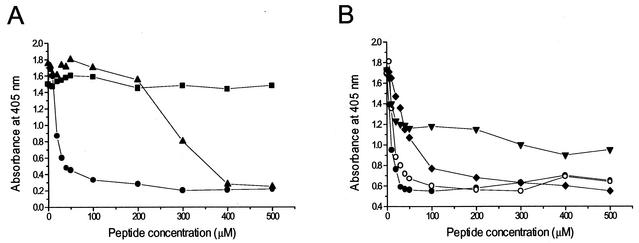
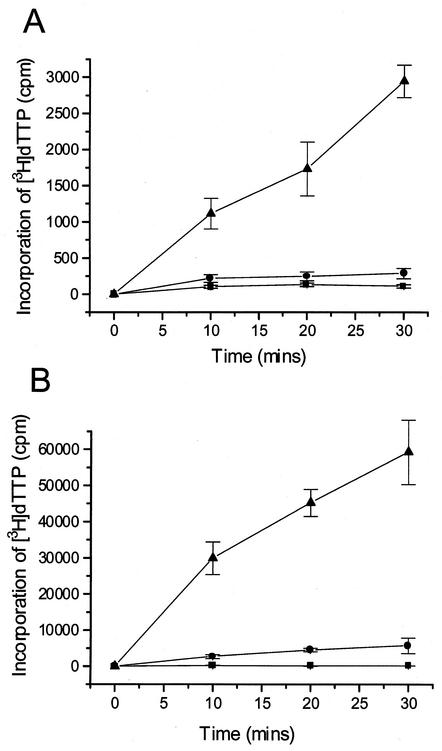
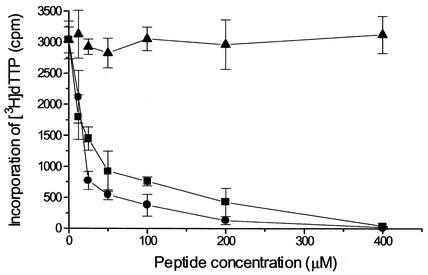
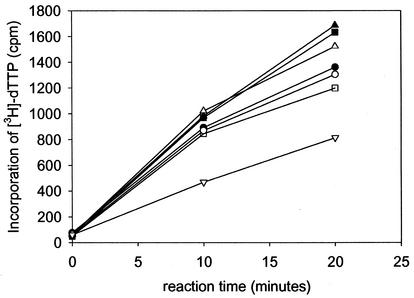
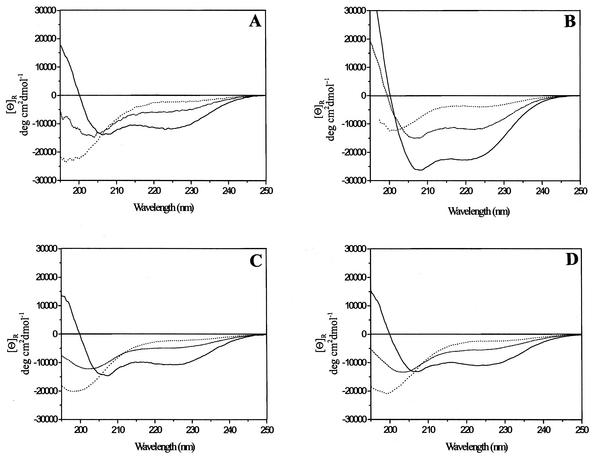
In common with other herpesviruses, the human cytomegalovirus (HCMV) DNA polymerase contains a catalytic subunit (Pol or UL54) and an accessory protein (UL44) that is thought to increase the processivity of the enzyme. The observation that antisense inhibition of UL44 synthesis in HCMV-infected cells strongly inhibits viral DNA replication, together with the structural similarity predicted for the herpesvirus processivity subunits, highlights the importance of the accessory protein for virus growth and raises the possibility that the UL54/UL44 interaction might be a valid target for antiviral drugs. To investigate this possibility, overlapping peptides spanning residues 1161 to 1242 of UL54 were synthesized and tested for inhibition of the interaction between purified UL54 and UL44 proteins. A peptide, LPRRLHLEPAFLPYSVKAHECC, corresponding to residues 1221 to 1242 at the very C terminus of UL54, disrupted both the physical interaction between the two proteins and specifically inhibited the stimulation of UL54 by UL44. A mutant peptide lacking the two carboxy-terminal cysteines was markedly less inhibitory, suggesting a role for these residues in the UL54/UL44 interaction. Circular dichroism spectroscopy indicated that the UL54 C-terminal peptide can adopt a partially alpha-helical structure. Taken together, these results indicate that the two subunits of HCMV DNA polymerase most likely interact in a way which is analogous to that of the two subunits of herpes simplex virus DNA polymerase, even though there is no sequence homology in the binding site, and suggest that the UL54 peptide, or derivatives thereof, could form the basis for developing a new class of anti-HCMV inhibitors that act by disrupting the UL54/UL44 interaction.
Figures
Similar articles
-
Residues of human cytomegalovirus DNA polymerase catalytic subunit UL54 that are necessary and sufficient for interaction with the accessory protein UL44.J Virol. 2004 Jan;78(1):158-67. doi: 10.1128/jvi.78.1.158-167.2004. J Virol. 2004. PMID: 14671097 Free PMC article.
-
Specific residues in the connector loop of the human cytomegalovirus DNA polymerase accessory protein UL44 are crucial for interaction with the UL54 catalytic subunit.J Virol. 2004 Sep;78(17):9084-92. doi: 10.1128/JVI.78.17.9084-9092.2004. J Virol. 2004. PMID: 15308704 Free PMC article.
-
Crystal structure of the cytomegalovirus DNA polymerase subunit UL44 in complex with the C terminus from the catalytic subunit. Differences in structure and function relative to unliganded UL44.J Biol Chem. 2006 Feb 24;281(8):5224-32. doi: 10.1074/jbc.M506900200. Epub 2005 Dec 20. J Biol Chem. 2006. PMID: 16371349
-
Herpesvirus DNA polymerases: Structures, functions and inhibitors.Virus Res. 2017 Apr 15;234:177-192. doi: 10.1016/j.virusres.2017.01.019. Epub 2017 Jan 30. Virus Res. 2017. PMID: 28153606 Review.
-
Herpesvirus DNA polymerase processivity factors: Not just for DNA synthesis.Virus Res. 2021 Jun;298:198394. doi: 10.1016/j.virusres.2021.198394. Epub 2021 Mar 26. Virus Res. 2021. PMID: 33775751 Review.
Cited by
-
The crystal structure of PF-8, the DNA polymerase accessory subunit from Kaposi's sarcoma-associated herpesvirus.J Virol. 2009 Dec;83(23):12215-28. doi: 10.1128/JVI.01158-09. Epub 2009 Sep 16. J Virol. 2009. PMID: 19759157 Free PMC article.
-
Small molecule inhibitors of influenza A and B viruses that act by disrupting subunit interactions of the viral polymerase.Proc Natl Acad Sci U S A. 2012 Apr 17;109(16):6247-52. doi: 10.1073/pnas.1119817109. Epub 2012 Apr 2. Proc Natl Acad Sci U S A. 2012. PMID: 22474359 Free PMC article.
-
Live-Cell Analysis of Human Cytomegalovirus DNA Polymerase Holoenzyme Assembly by Resonance Energy Transfer Methods.Microorganisms. 2021 Apr 26;9(5):928. doi: 10.3390/microorganisms9050928. Microorganisms. 2021. PMID: 33925913 Free PMC article.
-
The 6-aminoquinolone WC5 inhibits human cytomegalovirus replication at an early stage by interfering with the transactivating activity of viral immediate-early 2 protein.Antimicrob Agents Chemother. 2010 May;54(5):1930-40. doi: 10.1128/AAC.01730-09. Epub 2010 Mar 1. Antimicrob Agents Chemother. 2010. PMID: 20194695 Free PMC article.
-
Cloning, expression, and functional characterization of the equine herpesvirus 1 DNA polymerase and its accessory subunit.J Virol. 2006 Jul;80(13):6247-58. doi: 10.1128/JVI.02551-05. J Virol. 2006. PMID: 16775312 Free PMC article.
References
-
- Bishop, D. H. 1992. Baculovirus expression vectors. Semin. Virol. 3:253-264.
-
- Bohm, G., R. Muhr, and R. Jaenicke. 1992. Quantitative analysis of protein far UV circular dichroism spectra by neural networks. Protein Eng. 5:191-195. - PubMed
-
- Bridges, K. G., Q. Hua, M. R. Brigham-Burke, J. D. Martin, P. Hensley, C. E. Dahl, P. Digard, M. A. Weiss, and D. M. Coen. 2000. Secondary structure and structure-activity relationships of peptides corresponding to the subunit interface of herpes simplex virus DNA polymerase. J. Biol. Chem. 275:472-478. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
