

A heterologous coiled coil can substitute for helix I of the Sindbis virus capsid protein
- PMID: 12857904
- PMCID: PMC165231
- DOI: 10.1128/jvi.77.15.8345-8353.2003
A heterologous coiled coil can substitute for helix I of the Sindbis virus capsid protein
Abstract
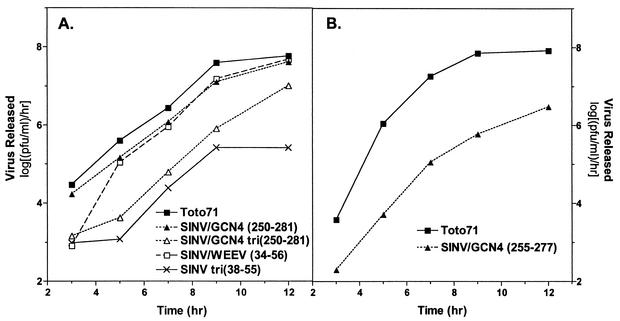
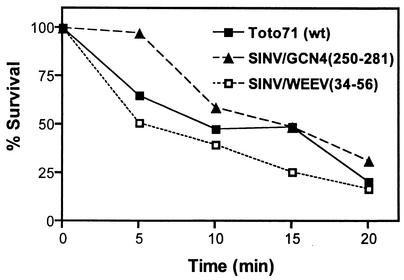
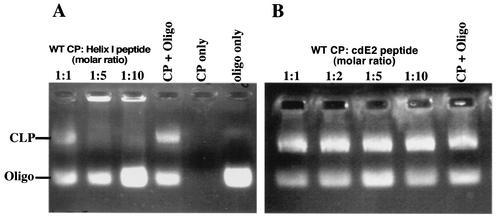
Alphavirus core assembly proceeds along an assembly pathway involving a dimeric assembly intermediate. Several regions of the alphavirus capsid protein have been implicated in promoting and stabilizing this dimerization, including a putative heptad repeat sequence named helix I. This sequence, which spans residues 38 to 55 of the Sindbis virus capsid protein, was implicated in stabilizing dimeric contacts initiated through the C-terminal two-thirds of the capsid protein and nucleic acid. The studies presented here demonstrate that helix I can be functionally replaced by the corresponding sequence of a related alphavirus, western equine encephalitis virus, and also by an unrelated sequence from the yeast transcription activator, GCN4, that was previously shown to form a dimeric coiled coil. Replacing helix I with the entire leucine zipper domain of GCN4 (residues 250 to 281) produced a virus with the wild-type phenotype as determined by plaque assay and one-step growth analysis. However, replacement of helix I with a GCN4 sequence that favored trimer formation produced a virus that exhibited approximately 40-fold reduction in virus replication compared to the wild-type Sindbis virus. Changing residues within the Sindbis virus helix I sequence to favor trimer formation also produced a virus with reduced replication. Peptides corresponding to helix I inhibited core-like particle assembly in vitro. On the basis of these studies, it is proposed that helix I favors capsid protein-capsid protein interactions through the formation of dimeric coiled-coil interactions and may stabilize assembly intermediates in the alphavirus nucleocapsid core assembly pathway.
Figures

Similar articles
-
Alphavirus capsid protein helix I controls a checkpoint in nucleocapsid core assembly.J Virol. 2006 Sep;80(18):8848-55. doi: 10.1128/JVI.00619-06. J Virol. 2006. PMID: 16940497 Free PMC article.
-
Alphavirus nucleocapsid protein contains a putative coiled coil alpha-helix important for core assembly.J Virol. 2001 Jan;75(1):1-10. doi: 10.1128/JVI.75.1.1-10.2001. J Virol. 2001. PMID: 11119567 Free PMC article.
-
The SD1 Subdomain of Venezuelan Equine Encephalitis Virus Capsid Protein Plays a Critical Role in Nucleocapsid and Particle Assembly.J Virol. 2015 Dec 9;90(4):2008-20. doi: 10.1128/JVI.02680-15. Print 2016 Feb 15. J Virol. 2015. PMID: 26656680 Free PMC article.
-
The sticky business of Alphavirus capsid-host interactions.Trends Microbiol. 2025 Mar;33(3):321-339. doi: 10.1016/j.tim.2024.11.003. Epub 2024 Dec 10. Trends Microbiol. 2025. PMID: 39665907 Review.
-
Are weak protein-protein interactions the general rule in capsid assembly?Virology. 2003 Oct 25;315(2):269-74. doi: 10.1016/s0042-6822(03)00586-5. Virology. 2003. PMID: 14585329 Review. No abstract available.
Cited by
-
A novel system for visualizing alphavirus assembly.J Virol Methods. 2015 Sep 15;222:158-63. doi: 10.1016/j.jviromet.2015.06.013. Epub 2015 Jun 27. J Virol Methods. 2015. PMID: 26122073 Free PMC article.
-
Adaptive changes in alphavirus mRNA translation allowed colonization of vertebrate hosts.J Virol. 2012 Sep;86(17):9484-94. doi: 10.1128/JVI.01114-12. Epub 2012 Jul 3. J Virol. 2012. PMID: 22761388 Free PMC article.
-
Capsid-E2 Interactions Rescue Core Assembly in Viruses That Cannot Form Cytoplasmic Nucleocapsid Cores.J Virol. 2021 Oct 27;95(22):e0106221. doi: 10.1128/JVI.01062-21. Epub 2021 Sep 8. J Virol. 2021. PMID: 34495691 Free PMC article.
-
Venezuelan Equine Encephalitis Virus Capsid-The Clever Caper.Viruses. 2017 Sep 29;9(10):279. doi: 10.3390/v9100279. Viruses. 2017. PMID: 28961161 Free PMC article. Review.
-
Venezuelan equine encephalitis virus capsid protein inhibits nuclear import in Mammalian but not in mosquito cells.J Virol. 2008 Apr;82(8):4028-41. doi: 10.1128/JVI.02330-07. Epub 2008 Feb 6. J Virol. 2008. PMID: 18256144 Free PMC article.
References
-
- Casjens, S. 1997. Principles of virion structure, function, and assembly, p. 3-37. In W. Chiu, R. M. Burnett, and R. Garcia (ed.), Structural biology of viruses. Oxford University Press, New York, N.Y.
-
- Choi, H.-K., L. Tong, W. Minor, P. Dumas, U. Boege, M. G. Rossmann, and G. Wengler. 1991. Structure of Sindbis virus core protein reveals a chymotrypsin-like serine proteinase and the organization of the virion. Nature 354:37-43. - PubMed
-
- Coombs, K., and D. T. Brown. 1987. Organization of the Sindbis virus nucleocapsid as revealed by bifunctional cross-linking agents. J. Mol. Biol. 195:359-371. - PubMed
-
- DeLano, W. L., and A. T. Brunger. 1994. Helix packing in proteins: prediction and energetic analysis of dimeric, trimeric, and tetrameric GCN4 coiled coil structures. Proteins 20:105-123. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
