

Role of alpha/beta interferons in the attenuation and immunogenicity of recombinant bovine respiratory syncytial viruses lacking NS proteins
- PMID: 12857912
- PMCID: PMC165239
- DOI: 10.1128/jvi.77.15.8426-8439.2003
Role of alpha/beta interferons in the attenuation and immunogenicity of recombinant bovine respiratory syncytial viruses lacking NS proteins
Abstract
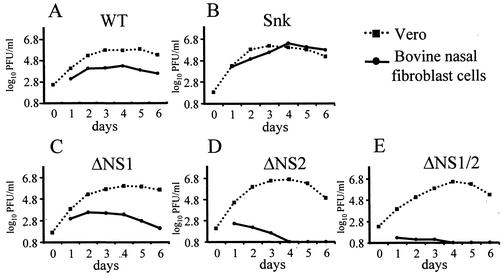
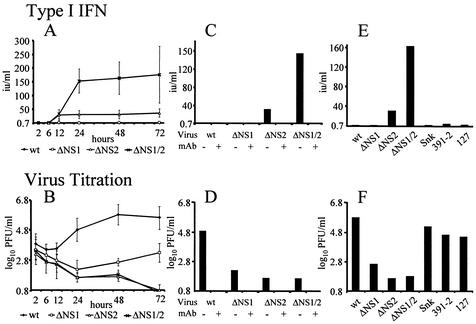
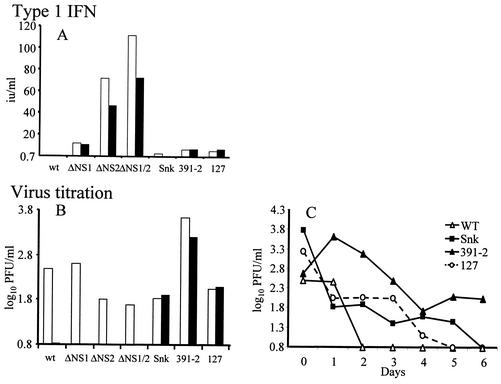
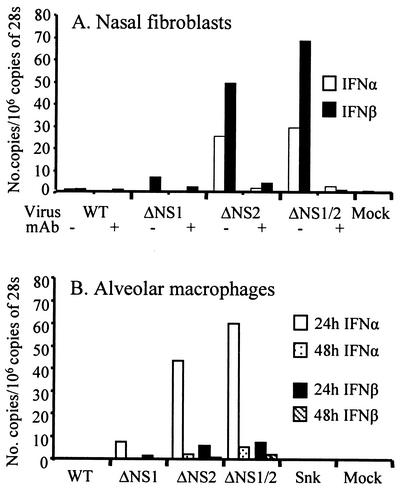
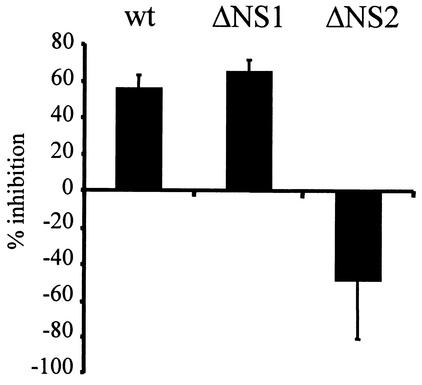
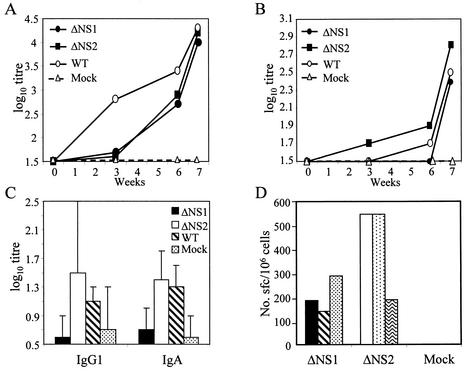
Alpha/beta interferons (IFN-alpha/beta) are not only a powerful first line of defense against pathogens but also have potent immunomodulatory activities. Many viruses have developed mechanisms of subverting the IFN system to enhance their virulence. Previous studies have demonstrated that the nonstructural (NS) genes of bovine respiratory syncytial virus (BRSV) counteract the antiviral effects of IFN-alpha/beta. Here we demonstrate that, in contrast to wild-type BRSVs, recombinant BRSVs (rBRSVs) lacking the NS proteins, and those lacking NS2 in particular, are strong inducers of IFN-alpha/beta in bovine nasal fibroblasts and bronchoalveolar macrophages. Furthermore, whereas the NS deletion mutants replicated to wild-type rBRSV levels in cells lacking a functional IFN-alpha/beta system, their replication was severely attenuated in IFN-competent cells and in young calves. These results suggest that the NS proteins block the induction of IFN-alpha/beta gene expression and thereby increase the virulence of BRSV. Despite their poor replication in the respiratory tract of young calves, prior infection with virus lacking either the NS1 or the NS2 protein induced serum antibodies and protection against challenge with virulent BRSV. The greater level of protection induced by the NS2, than by the NS1, deletion mutant, was associated with higher BRSV-specific antibody titers and greater priming of BRSV-specific, IFN-gamma-producing CD4(+) T cells. Since there were no detectable differences in the ability of these mutants to replicate in the bovine respiratory tract, the greater immunogenicity of the NS2 deletion mutant may be associated with the greater ability of this virus to induce IFN-alpha/beta.
Figures
Similar articles
-
Bovine respiratory syncytial virus nonstructural proteins NS1 and NS2 cooperatively antagonize alpha/beta interferon-induced antiviral response.J Virol. 2000 Sep;74(18):8234-42. doi: 10.1128/jvi.74.18.8234-8242.2000. J Virol. 2000. PMID: 10954520 Free PMC article.
-
Respiratory syncytial virus (RSV) nonstructural (NS) proteins as host range determinants: a chimeric bovine RSV with NS genes from human RSV is attenuated in interferon-competent bovine cells.J Virol. 2002 May;76(9):4287-93. doi: 10.1128/jvi.76.9.4287-4293.2002. J Virol. 2002. PMID: 11932394 Free PMC article.
-
Nonstructural proteins NS1 and NS2 of bovine respiratory syncytial virus block activation of interferon regulatory factor 3.J Virol. 2003 Aug;77(16):8661-8. doi: 10.1128/jvi.77.16.8661-8668.2003. J Virol. 2003. PMID: 12885884 Free PMC article.
-
[Immunobiology of bovine respiratory syncytial virus infections].Tijdschr Diergeneeskd. 1998 Nov 15;123(22):658-62. Tijdschr Diergeneeskd. 1998. PMID: 9836385 Review. Dutch.
-
Bovine respiratory syncytial virus infection: immunopathogenic mechanisms.Anim Health Res Rev. 2007 Dec;8(2):207-13. doi: 10.1017/S1466252307001405. Anim Health Res Rev. 2007. PMID: 18218161 Review.
Cited by
-
Neonatal calf infection with respiratory syncytial virus: drawing parallels to the disease in human infants.Viruses. 2012 Dec;4(12):3731-53. doi: 10.3390/v4123731. Viruses. 2012. PMID: 23342375 Free PMC article. Review.
-
Transcriptional activation of alpha/beta interferon genes: interference by nonsegmented negative-strand RNA viruses.J Virol. 2005 May;79(9):5241-8. doi: 10.1128/JVI.79.9.5241-5248.2005. J Virol. 2005. PMID: 15827138 Free PMC article. Review. No abstract available.
-
Inhibition of toll-like receptor 7- and 9-mediated alpha/beta interferon production in human plasmacytoid dendritic cells by respiratory syncytial virus and measles virus.J Virol. 2005 May;79(9):5507-15. doi: 10.1128/JVI.79.9.5507-5515.2005. J Virol. 2005. PMID: 15827165 Free PMC article.
-
Immune response in the absence of neurovirulence in mice infected with m protein mutant vesicular stomatitis virus.J Virol. 2008 Sep;82(18):9273-7. doi: 10.1128/JVI.00915-08. Epub 2008 Jul 9. J Virol. 2008. PMID: 18614644 Free PMC article.
-
Reprogramming viral immune evasion for a rational design of next-generation vaccines for RNA viruses.Front Immunol. 2023 Apr 17;14:1172000. doi: 10.3389/fimmu.2023.1172000. eCollection 2023. Front Immunol. 2023. PMID: 37138878 Free PMC article. Review.
References
-
- Bell, D. M., N. J. Roberts, and C. B. Hall. 1983. Different antiviral spectra of human macrophage interferon activities. Nature 305:319-321. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
