

CD4+ T cell polarization in mice is modulated by strain-specific major histocompatibility complex-independent differences within dendritic cells
- PMID: 12860929
- PMCID: PMC2194066
- DOI: 10.1084/jem.20021893
CD4+ T cell polarization in mice is modulated by strain-specific major histocompatibility complex-independent differences within dendritic cells
Abstract
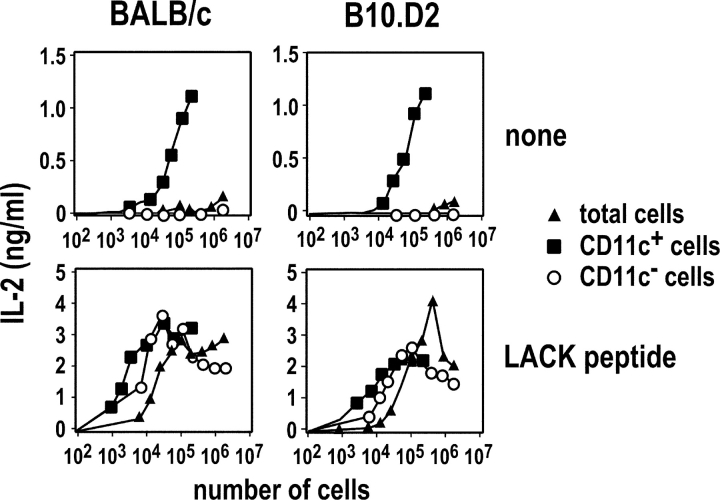
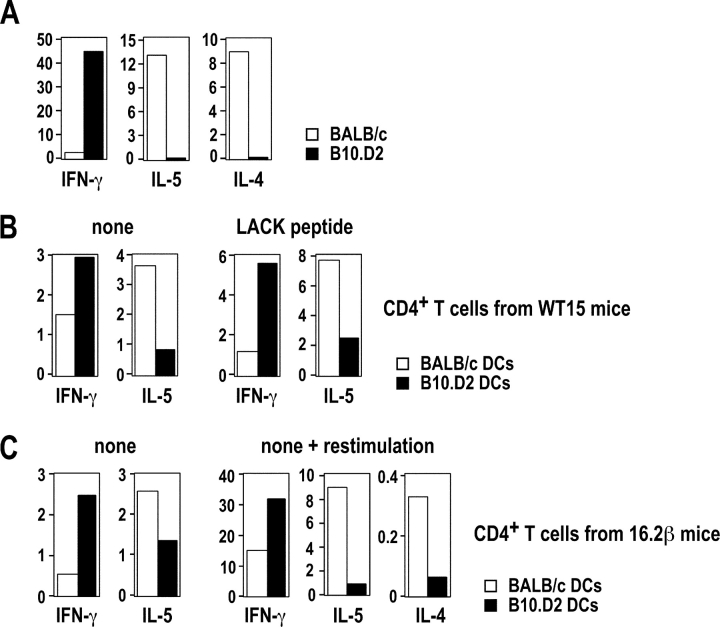
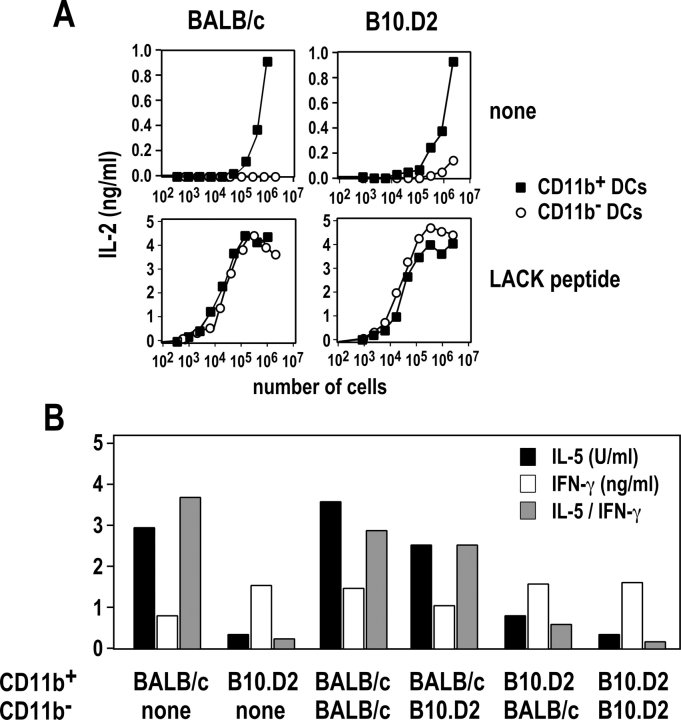
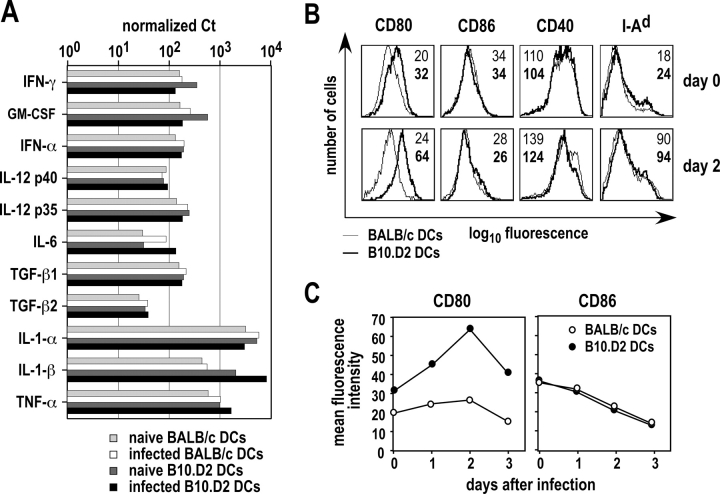
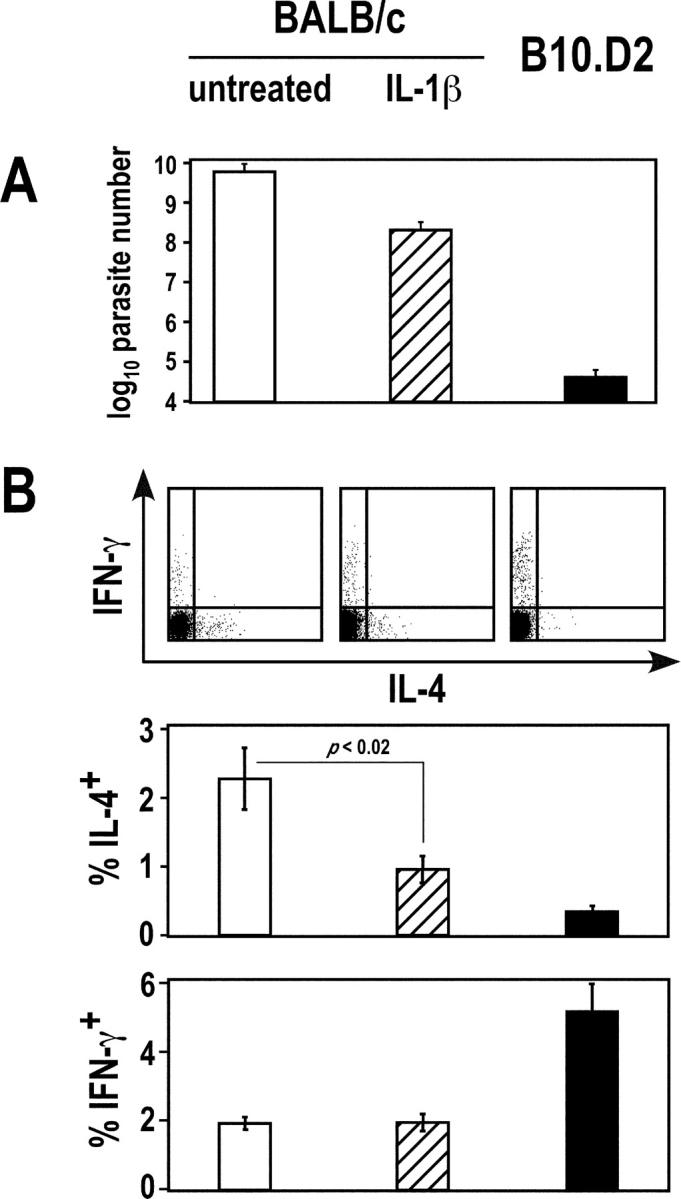
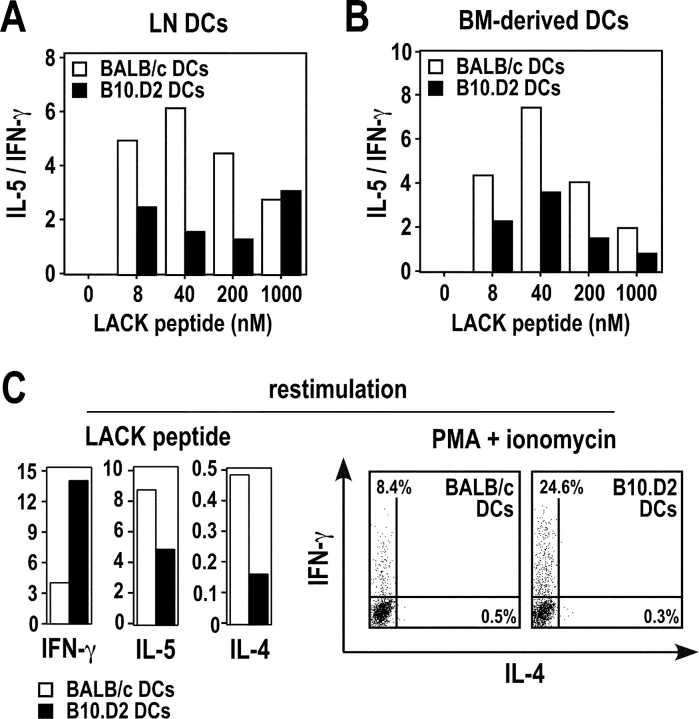
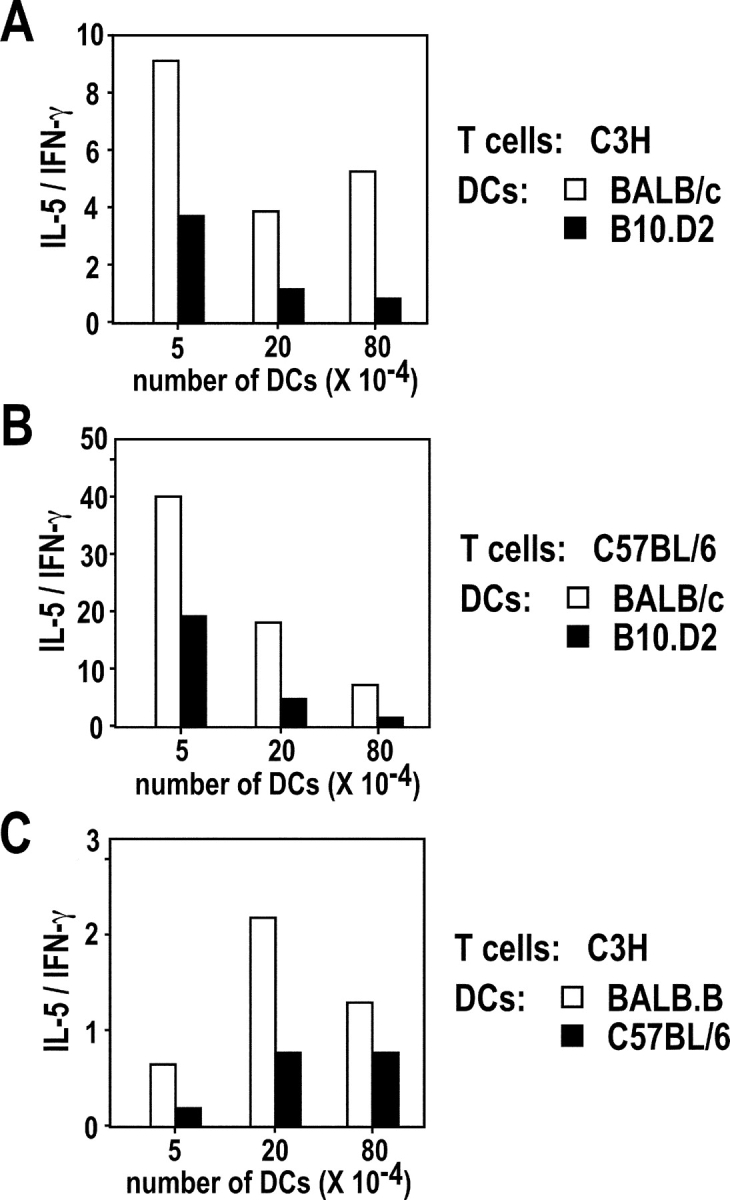
Resistance and susceptibility to Leishmania major in mice are determined by multiple genes and correlate with the preferential development of Th1 and Th2 responses, respectively. Here, we found that CD11b+ dendritic cells (DCs) prime parasite-specific CD4+ T cells in both susceptible BALB/c (H2-d) and resistant B10.D2 (H2-d) mice. However, BALB/c and B10.D2 DCs from L. major-infected mice differ in their ability to polarize naive T cells into Th1 or Th2 effector cells. This difference is cell-intrinsic, is not restricted to H2-d mice, and is observed with both parasite-specific and allospecific CD4+ T cells. Thus, strain-specific differences within CD11b+ DCs influence the ability of inbred mice to mount polarized CD4+ T cell responses.
Figures
Comment in
-
The importance of CD11b+ dendritic cells in CD4+ T cell activation in vivo: with help from interleukin 1.J Exp Med. 2003 Jul 21;198(2):185-90. doi: 10.1084/jem.20030737. Epub 2003 Jul 14. J Exp Med. 2003. PMID: 12860934 Free PMC article. Review. No abstract available.
References
-
- Reiner, S.L., and R.M. Locksley. 1995. The regulation of immunity to Leishmania major. Annu. Rev. Immunol. 13:151–177. - PubMed
-
- Beebe, A.M., S. Mauze, N.J. Schork, and R.L. Coffman. 1997. Serial backcross mapping of multiple loci associated with resistance to Leishmania major in mice. Immunity. 6:551–557. - PubMed
-
- Coffman, R.L., and A.M. Beebe. 1998. Genetic control of the T cell response to Leishmania major infection. Adv. Exp. Med. Biol. 452:61–66. - PubMed
-
- Roberts, L.J., T.M. Baldwin, T.P. Speed, E. Handman, and S.J. Foote. 1999. Chromosomes X, 9, and the H2 locus interact epistatically to control Leishmania major infection. Eur. J. Immunol. 29:3047–3050. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
