

Interleukin 1alpha promotes Th1 differentiation and inhibits disease progression in Leishmania major-susceptible BALB/c mice
- PMID: 12860932
- PMCID: PMC2194079
- DOI: 10.1084/jem.20030159
Interleukin 1alpha promotes Th1 differentiation and inhibits disease progression in Leishmania major-susceptible BALB/c mice
Abstract
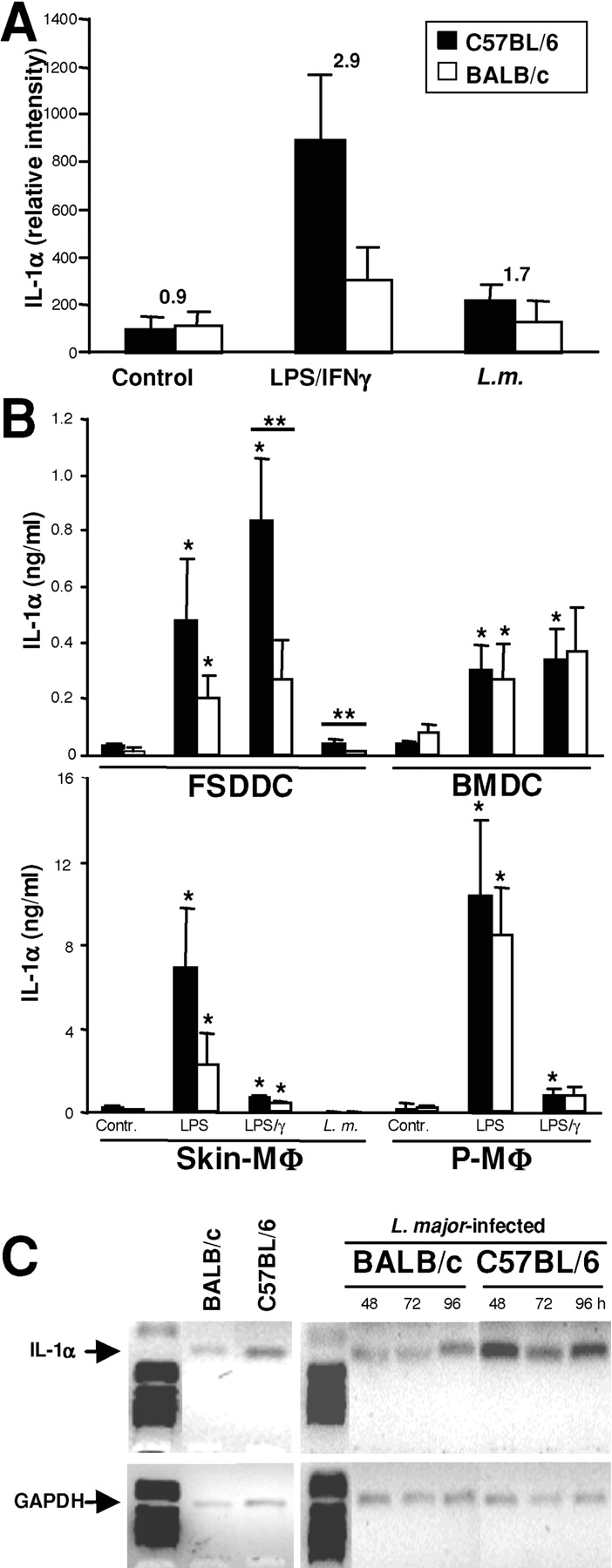
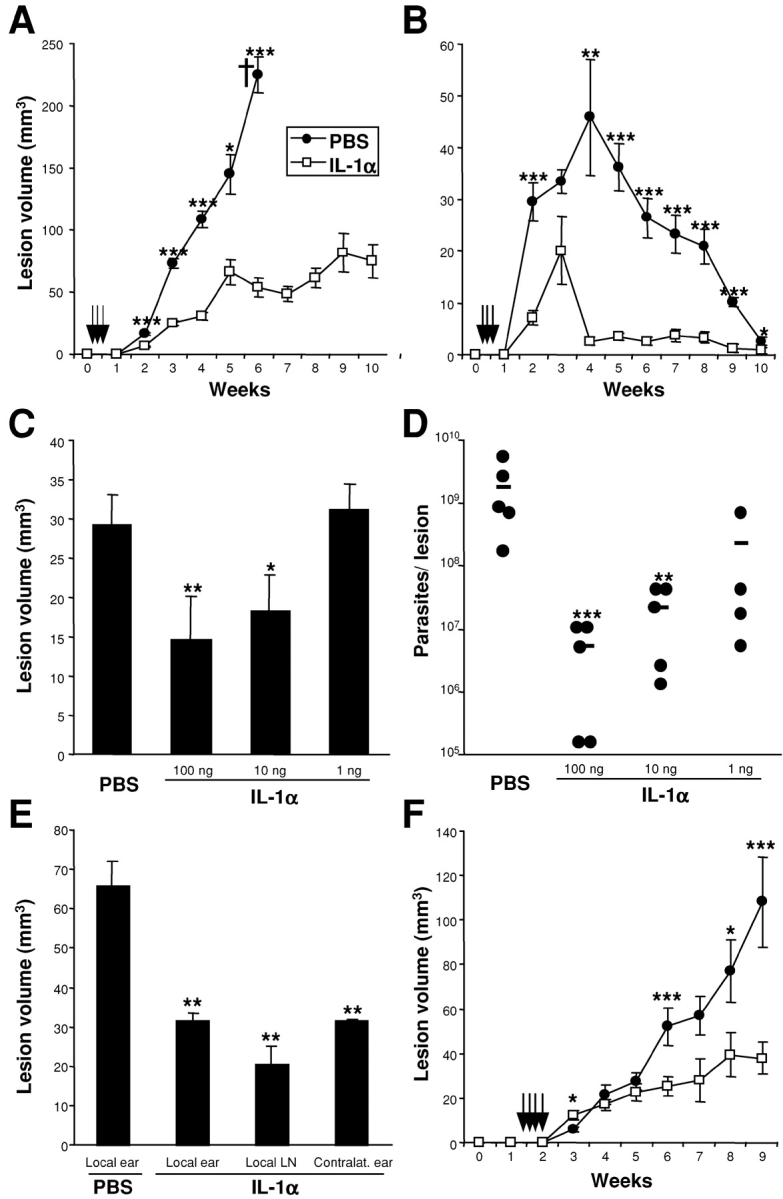
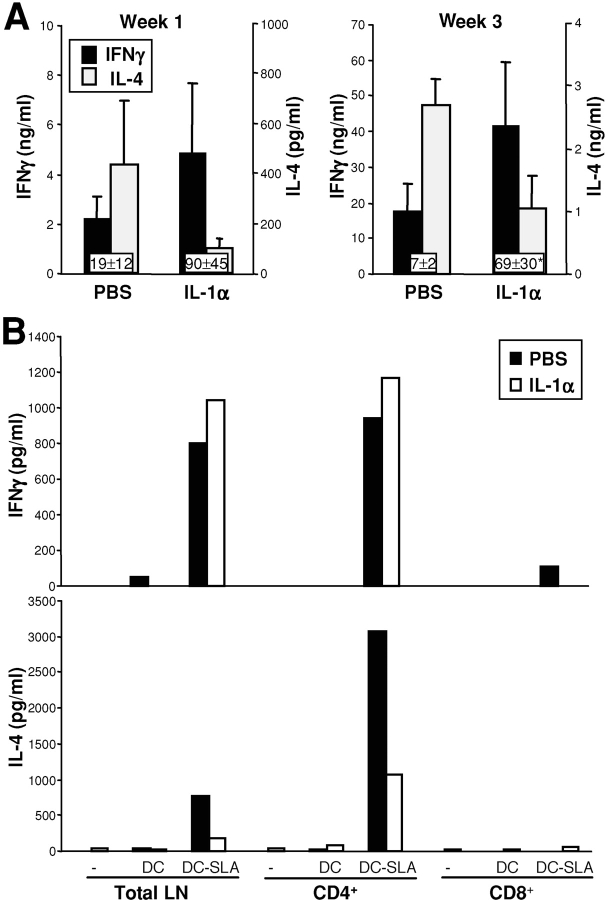
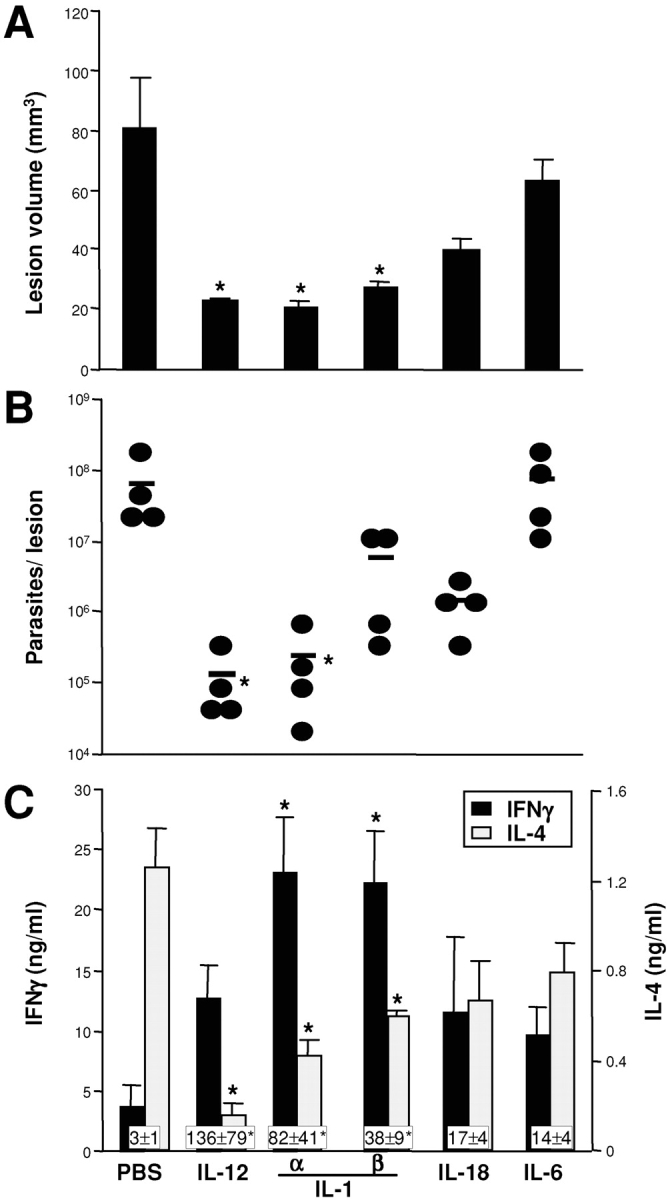
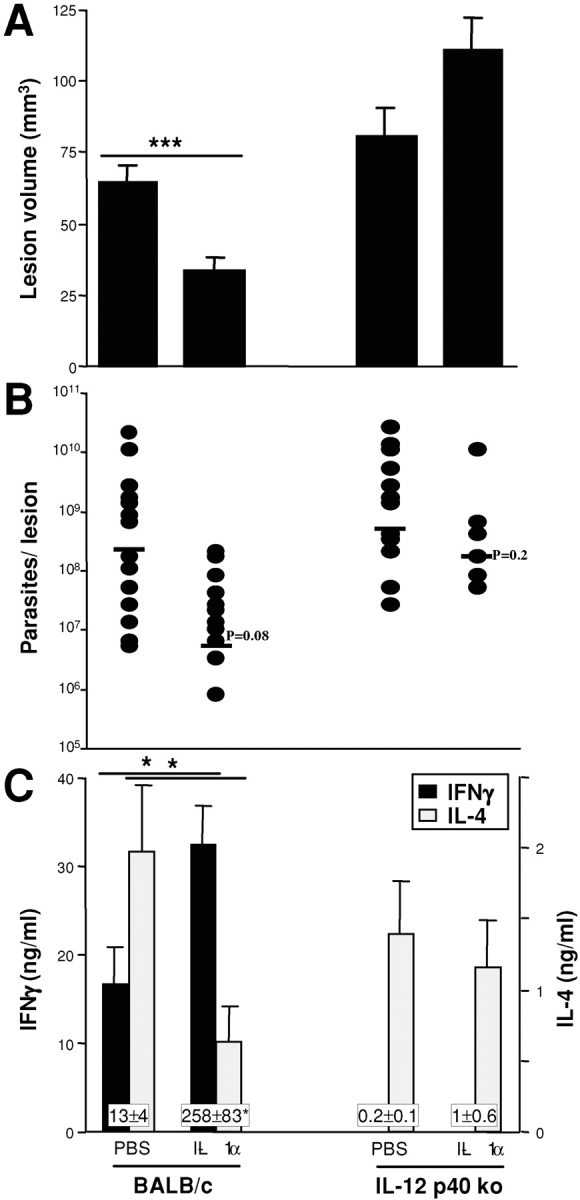
Protective immunity against pathogens such as Leishmania major is mediated by interleukin (IL)-12-dependent Th1-immunity. We have shown previously that skin-dendritic cells (DCs) from both resistant C57BL/6 and susceptible BALB/c mice release IL-12 when infected with L. major, and infected BALB/c DCs effectively vaccinate against leishmaniasis. To determine if cytokines other than IL-12 might influence disease outcome, we surveyed DCs from both strains for production of a variety of cytokines. Skin-DCs produced significantly less IL-1alpha in response to lipopolysaccharide/interferon gamma or L. major when expanded from BALB/c as compared with C57BL/6 mice. In addition, IL-1alpha mRNA accumulation in lymph nodes of L. major-infected BALB/c mice was approximately 3-fold lower than that in C57BL/6 mice. Local injections of IL-1alpha during the first 3 d after infection led to dramatic, persistent reductions in lesion sizes. In L. major-infected BALB/c mice, IL-1alpha administration resulted in increased Th1- and strikingly decreased Th2-cytokine production. IL-1alpha and IL-12 treatments were similarly effective, and IL-1alpha efficacy was strictly IL-12 dependent. These data indicate that transient local administration of IL-1alpha acts in conjunction with IL-12 to influence Th-development in cutaneous leishmaniasis and prevents disease progression in susceptible BALB/c mice, perhaps by enhancing DC-induced Th1-education. Differential production of IL-1 by C57BL/6 and BALB/c mice may provide a partial explanation for the disparate outcomes of infection in these mouse strains.
Figures
Comment in
-
The importance of CD11b+ dendritic cells in CD4+ T cell activation in vivo: with help from interleukin 1.J Exp Med. 2003 Jul 21;198(2):185-90. doi: 10.1084/jem.20030737. Epub 2003 Jul 14. J Exp Med. 2003. PMID: 12860934 Free PMC article. Review. No abstract available.
References
-
- Reiner, S.L., and R.M. Locksley. 1995. The regulation of immunity to Leishmania major. Annu. Rev. Immunol. 13:151–177. - PubMed
-
- Muller, I., T. Pedrazzini, P. Kropf, J. Louis, and G. Milon. 1991. Establishment of resistance to Leishmania major infection in susceptible BALB/c mice requires parasite-specific CD8+ T cells. Int. Immunol. 3:587–597. - PubMed
-
- Belkaid, Y., E. von Stebut, S. Mendez, R. Lira, E. Caler, S. Bertholet, M.C. Udey, and D.L. Sacks. 2002. CD8+ T cells are required for primary immunity in C57BL/6 mice following low-dose, intradermal challenge with Leishmania major. J. Immunol. 168:3992–4000. - PubMed
-
- Flohe, S.B., C. Bauer, S. Flohe, and H. Moll. 1998. Antigen-pulsed epidermal Langerhans cells protect susceptible mice from infection with the intracellular parasite Leishmania major. Eur. J. Immunol. 28:3800–3811. - PubMed
