

Rb and N-ras function together to control differentiation in the mouse
- PMID: 12861012
- PMCID: PMC165732
- DOI: 10.1128/MCB.23.15.5256-5268.2003
Rb and N-ras function together to control differentiation in the mouse
Abstract
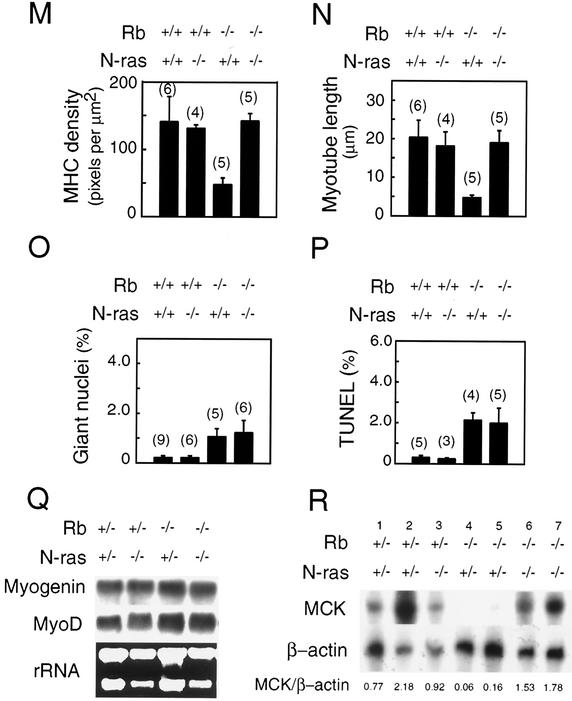
The product of the retinoblastoma tumor suppressor gene (Rb) can control cell proliferation and promote differentiation. Murine embryos nullizygous for Rb die midgestation with defects in cell cycle regulation, control of apoptosis, and terminal differentiation of several tissues, including skeletal muscle, nervous system, and lens. Previous cell culture-based experiments have suggested that the retinoblastoma protein (pRb) and Ras operate in a common pathway to control cellular differentiation. Here we have tested the hypothesis that the proto-oncogene N-ras participates in Rb-dependent regulation of differentiation by generating and characterizing murine embryos deficient in both N-ras and Rb. We show that deletion of N-ras rescues a unique subset of the developmental defects associated with nullizygosity of Rb, resulting in a significant extension of life span. Rb(-/-); N-ras(-/-) skeletal muscle has normal fiber density, myotube length and thickness, in contrast to Rb-deficient embryos. Additionally, Rb(-/-); N-ras(-/-) muscle shows a restoration in the expression of the late muscle-specific gene MCK, and this correlates with a significant potentiation of MyoD transcriptional activity in Rb(-/-); N-ras(-/-), compared to Rb(-/-) myoblasts in culture. The improved differentiation of skeletal muscle in Rb(-/-); N-ras(-/-) embryos occurs despite evidence of deregulated proliferation and apoptosis, as seen in Rb-deficient animals. Our findings suggest that the control of differentiation and proliferation by Rb are genetically separable.
Figures







References
-
- Bos, J. L. 1989. ras oncogenes in human cancer: a review. Cancer Res. 49:4682-4689. - PubMed
-
- Ceol, C. J., and H. R. Horvitz. 2001. dpl-1 DP and efl-1 E2F act with lin-35 Rb to antagonize Ras signaling in C. elegans vulval development. Mol. Cell 7:461-473. - PubMed
-
- Chen, P.-L., D. J. Riley, Y. Chen, and W.-H. Lee. 1996. Retinoblastoma protein positively regulates terminal adipocyte differentiation through direct interaction with C/EBPs. Genes Dev. 10:2794-2804. - PubMed
-
- Clarke, A. R., E. R. Maandag, M. van Roon, N. M. T. van der Lugt, M. van der Valk, M. L. Hooper, A. Berns, and H. te Riele. 1992. Requirement for a functional Rb-1 gene in murine development. Nature 359:328-330. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous