

Cdc34 self-association is facilitated by ubiquitin thiolester formation and is required for its catalytic activity
- PMID: 12861024
- PMCID: PMC165730
- DOI: 10.1128/MCB.23.15.5388-5400.2003
Cdc34 self-association is facilitated by ubiquitin thiolester formation and is required for its catalytic activity
Abstract
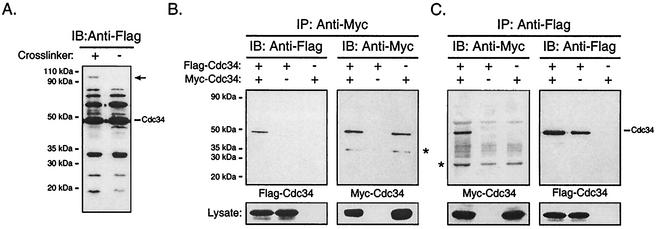
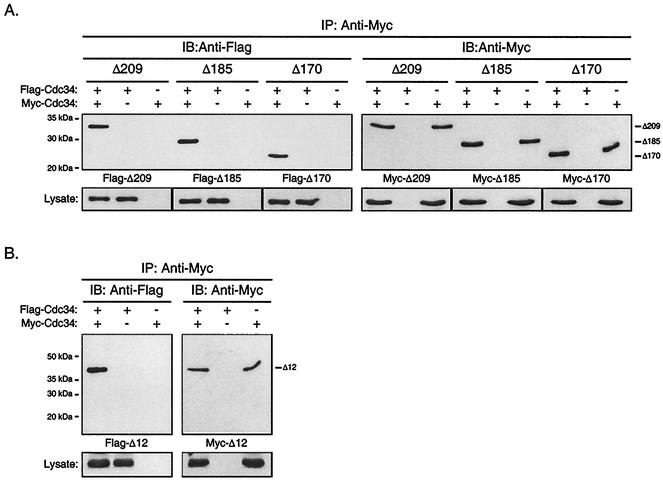
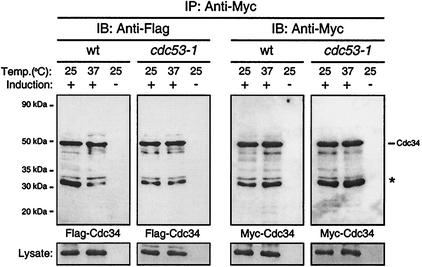
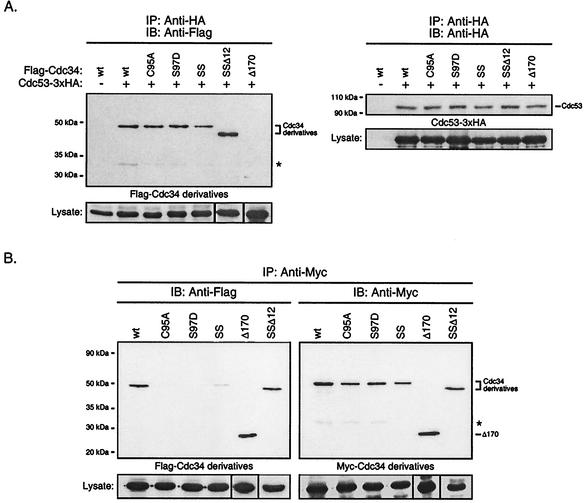
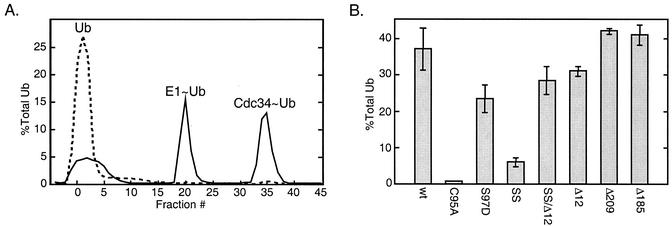
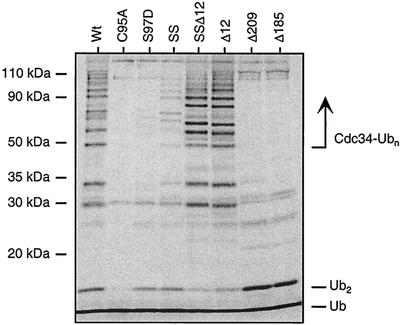
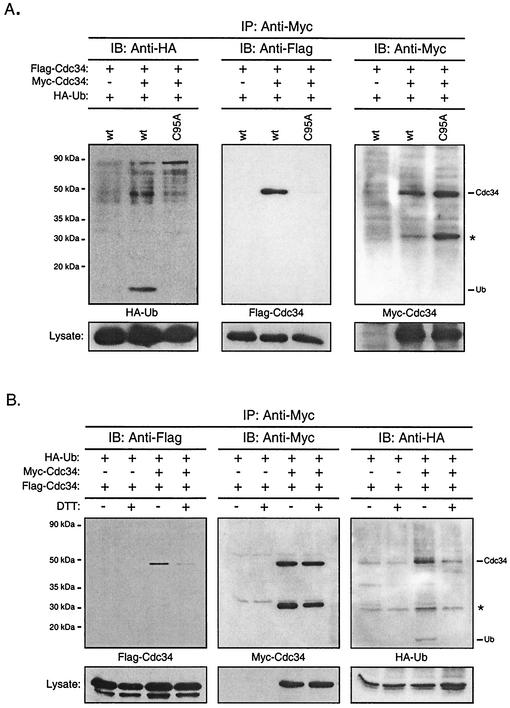
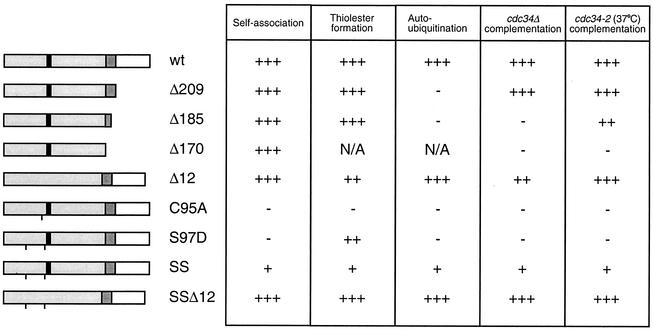
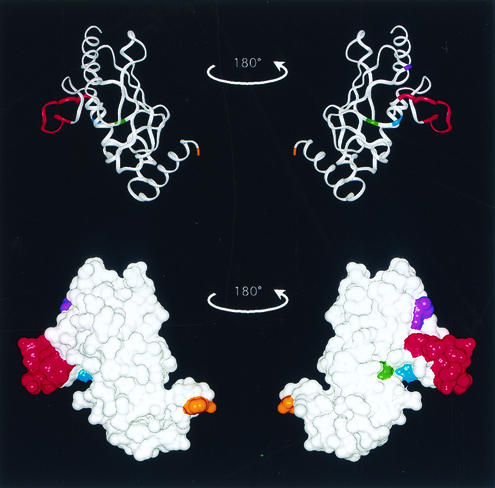
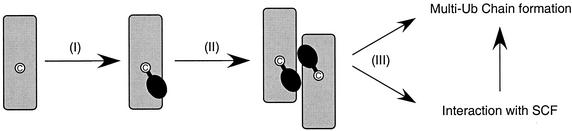
Using a coimmunoprecipitation strategy, we showed that the Cdc34 ubiquitin (Ub)-conjugating enzyme from Saccharomyces cerevisiae self-associates in cell lysates, thereby indicating an in vivo interaction. The ability of Cdc34 to interact with itself is not dependent on its association with the ubiquitin ligase Skp1-Cdc53/Cul1-Hrt1-F-box complex. Rather, this interaction depends upon the integrity of the Cdc34-Ub thiolester. Furthermore, several principal determinants within the Cdc34 catalytic domain, including the active-site cysteine, amino acid residues S73 and S97, and its catalytic domain insertion, also play a role in self-association. Mutational studies have shown that these determinants are functionally important in vivo and operate at the levels of both Cdc34-Ub thiolester formation and Cdc34-mediated multi-Ub chain assembly. These determinants are spatially situated in a region that is close to the active site, corresponding closely to the previously identified E2-Ub interface. These observations indicate that the formation of the Cdc34-Ub thiolester is important for Cdc34 self-association and that the interaction of Cdc34-Ub thiolesters is in turn a prerequisite for both multi-Ub chain assembly and Cdc34's essential function(s). A conclusion from these findings is that the placement of ubiquitin on the Cdc34 surface is a structurally important feature of Cdc34's function.
Figures
References
-
- Ausubel, F., R. Brent, R. E. Kingston, D. D. Moore, J. G. Seidman, J. A. Smith, and K. Struhl. 1995. Short protocols in molecular biology, 3rd ed. John Wiley & Sons, Inc., New York, N.Y.
-
- Banerjee, A., R. J. Deshaies, and V. Chau. 1995. Characterization of a dominant negative mutant of the cell cycle ubiquitin-conjugating enzyme Cdc34. J. Biol. Chem. 270:26209-26215. - PubMed
-
- Banerjee, A., L. Gregori, Y. Xu, and V. Chau. 1993. The bacterially expressed yeast CDC34 gene product can undergo autoubiquitination to form a multiubiquitin chain-linked protein. J. Biol. Chem. 268:5668-5675. - PubMed
-
- Chen, P., P. Johnson, T. Sommer, S. Jentsch, and M. Hochstrasser. 1993. Multiple ubiquitin-conjugating enzymes participate in the in vivo degradation of the yeast MAT alpha 2 repressor. Cell 74:357-369. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials