

CsrA regulates translation of the Escherichia coli carbon starvation gene, cstA, by blocking ribosome access to the cstA transcript
- PMID: 12867454
- PMCID: PMC165747
- DOI: 10.1128/JB.185.15.4450-4460.2003
CsrA regulates translation of the Escherichia coli carbon starvation gene, cstA, by blocking ribosome access to the cstA transcript
Abstract
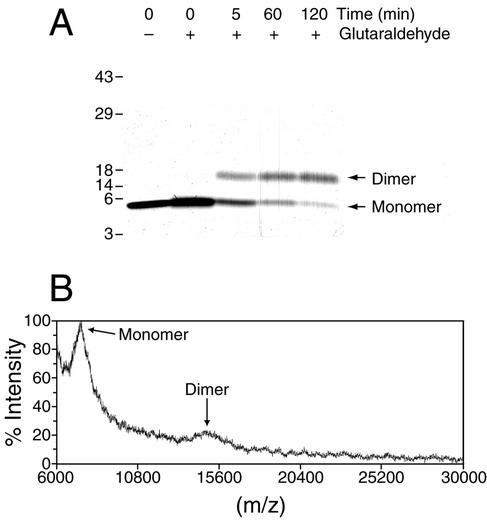
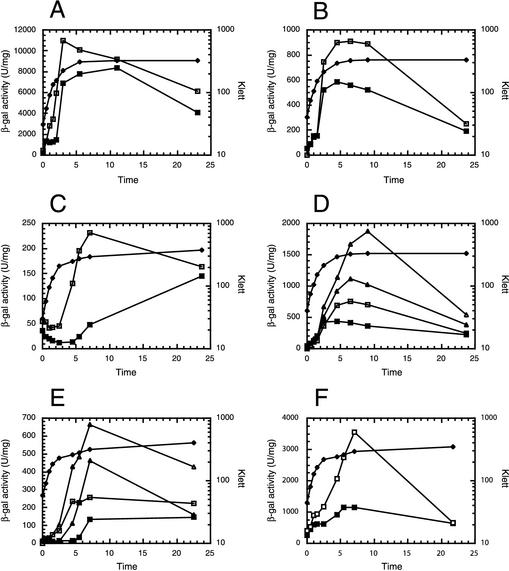
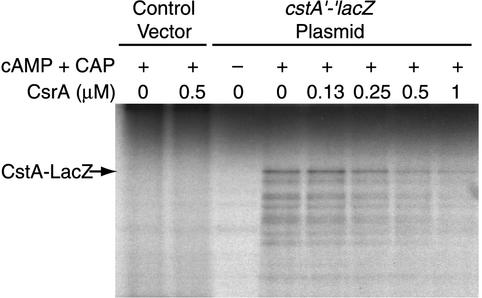
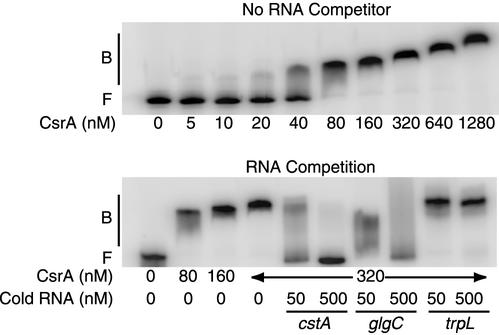
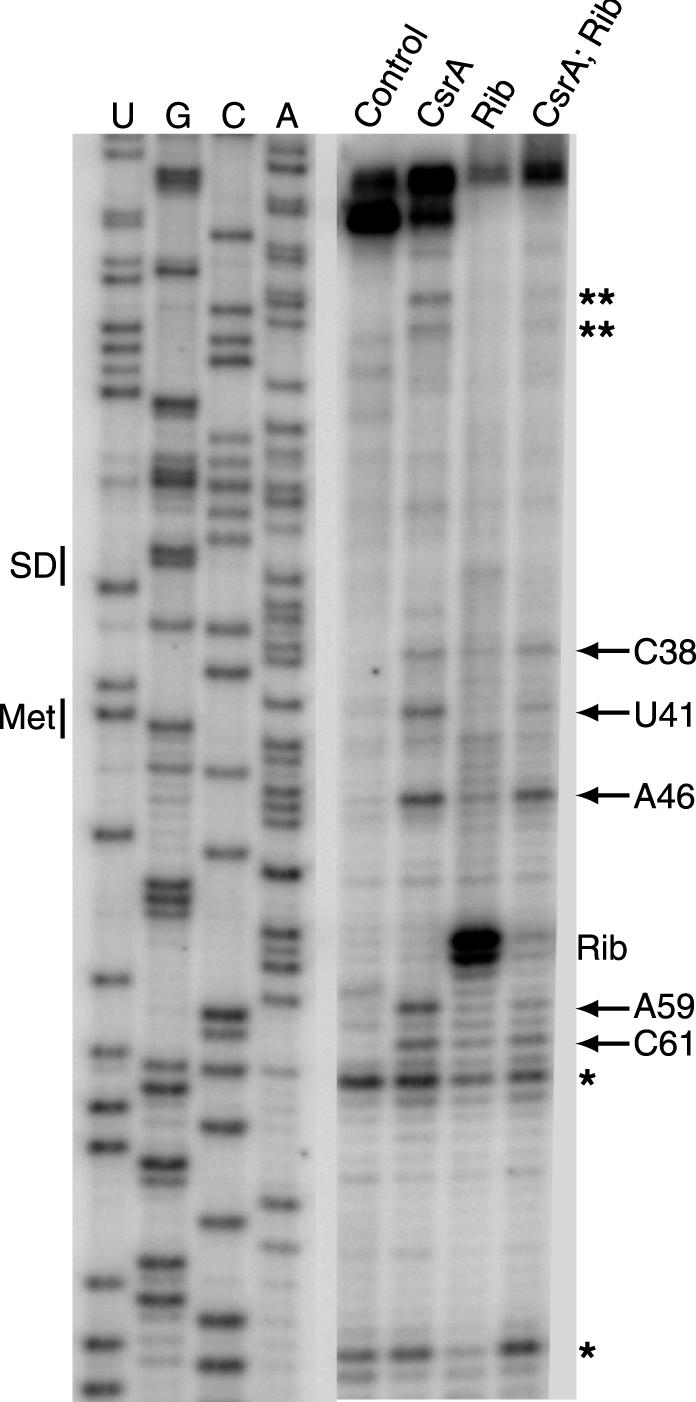
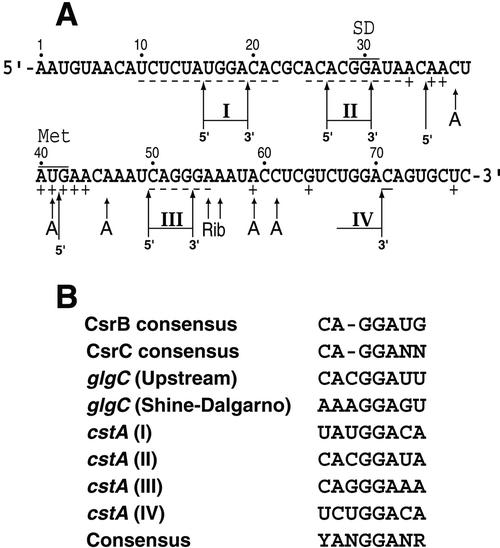
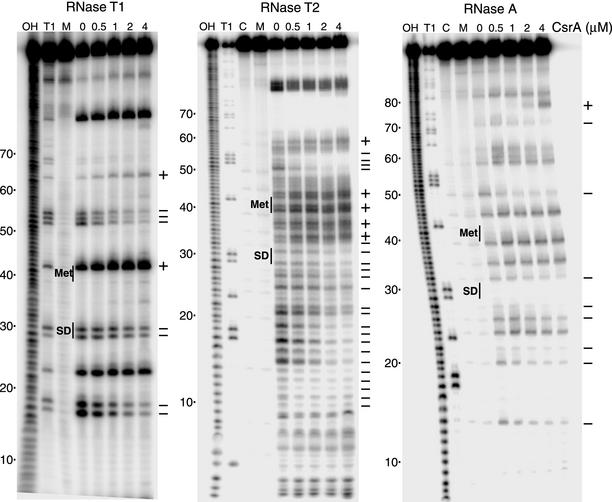
CsrA is a global regulator that binds to two sites in the glgCAP leader transcript, thereby blocking ribosome access to the glgC Shine-Dalgarno sequence. The upstream CsrA binding site (GCACACGGAU) was used to search the Escherichia coli genomic sequence for other genes that might be regulated by CsrA. cstA contained an exact match that overlapped its Shine-Dalgarno sequence. cstA was previously shown to be induced by carbon starvation and to encode a peptide transporter. Expression of a cstA'-'lacZ translational fusion in wild-type and csrA mutant strains was examined. Expression levels in the csrA mutant were approximately twofold higher when cells were grown in Luria broth (LB) and 5- to 10-fold higher when LB was supplemented with glucose. It was previously shown that cstA is regulated by the cyclic AMP (cAMP)-cAMP receptor protein complex and transcribed by Esigma(70). We investigated the influence of sigma(S) on cstA expression and found that a sigma(S) deficiency resulted in a threefold increase in cstA expression in wild-type and csrA mutant strains; however, CsrA-dependent regulation was retained. The mechanism of CsrA-mediated cstA regulation was also examined in vitro. Cross-linking studies demonstrated that CsrA is a homodimer. Gel mobility shift results showed that CsrA binds specifically to cstA RNA, while coupled-transcription-translation and toeprint studies demonstrated that CsrA regulates CstA synthesis by inhibiting ribosome binding to cstA transcripts. RNA footprint and boundary analyses revealed three or four CsrA binding sites, one of which overlaps the cstA Shine-Dalgarno sequence, as predicted. These results establish that CsrA regulates translation of cstA by sterically interfering with ribosome binding.
Figures

References
-
- Babitzke, P., J. T. Stults, S. J. Shire, and C. Yanofsky. 1994. TRAP, the trp RNA-binding attenuation protein of Bacillus subtilis, is a multisubunit complex that appears to recognize G/UAG repeats in the trpEDCFBA and trpG transcripts. J. Biol. Chem. 269:16597-16604. - PubMed
-
- Baker, C. S., I. Morozov, K. Suzuki, T. Romeo, and P. Babitzke. 2002. CsrA regulates glycogen biosynthesis by preventing translation of glgC in Escherichia coli. Mol. Microbiol. 44:1599-1610. - PubMed
-
- Bevilacqua, J. M., and P. C. Bevilacqua. 1998. Thermodynamic analysis of an RNA combinatorial library contained in a short hairpin. Biochemistry 37:15877-15884. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
