

Critical role for microglial NADPH oxidase in rotenone-induced degeneration of dopaminergic neurons
- PMID: 12867501
- PMCID: PMC6740554
- DOI: 10.1523/JNEUROSCI.23-15-06181.2003
Critical role for microglial NADPH oxidase in rotenone-induced degeneration of dopaminergic neurons
Abstract
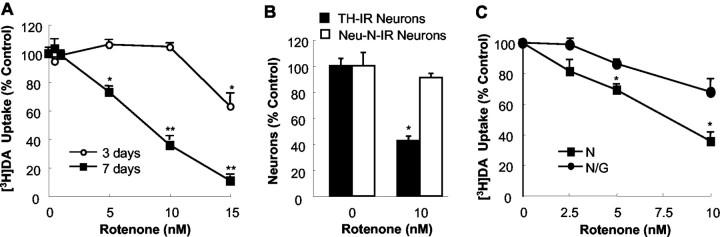
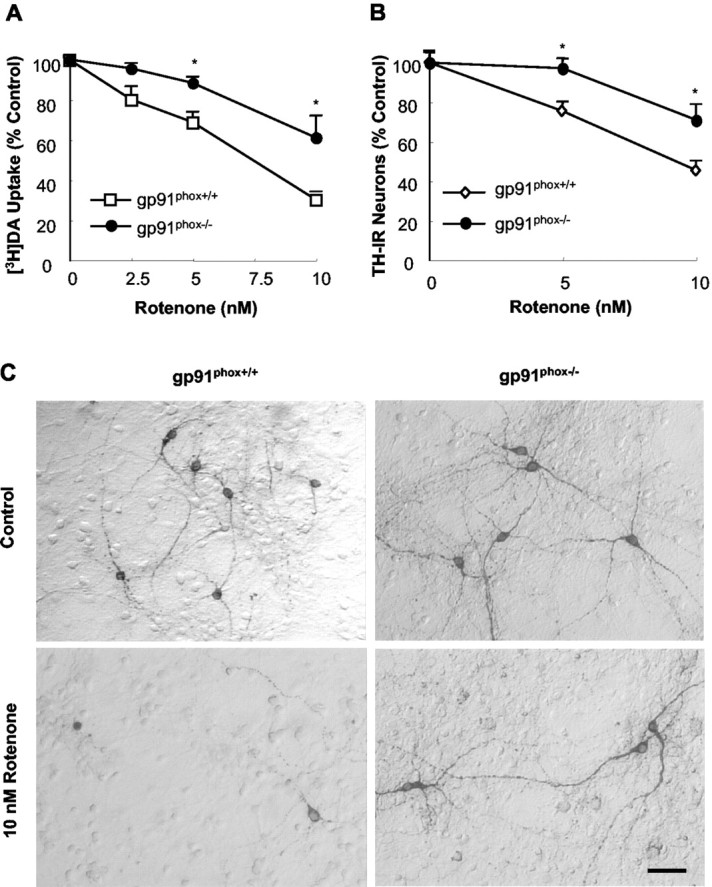
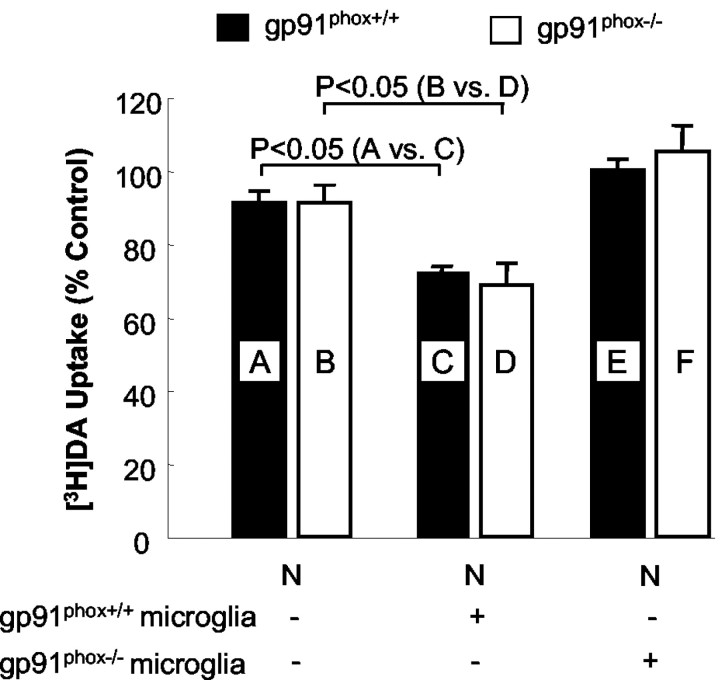
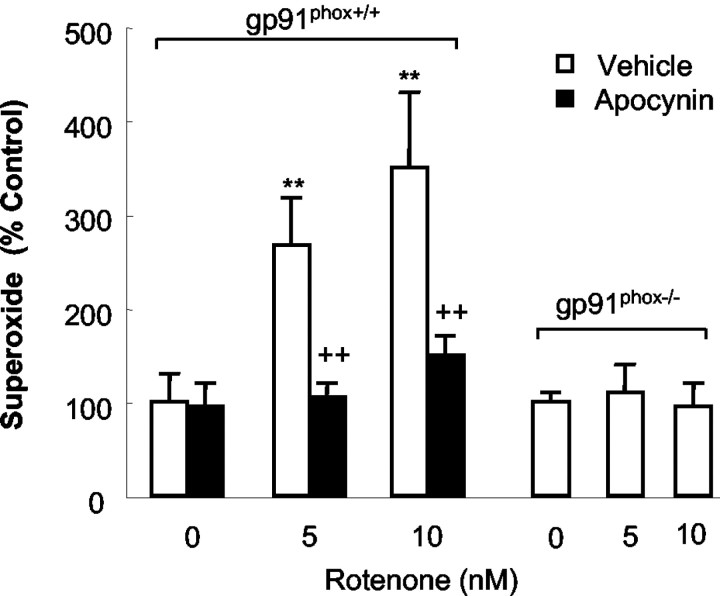
Increasing evidence has suggested an important role for environmental toxins such as pesticides in the pathogenesis of Parkinson's disease (PD). Chronic exposure to rotenone, a common herbicide, reproduces features of Parkinsonism in rats. Mechanistically, rotenone-induced dopaminergic neurodegeneration has been associated with both its inhibition of neuronal mitochondrial complex I and the enhancement of activated microglia. Our previous studies with NADPH oxidase inhibitors, diphenylene iodonium and apocynin, suggested that NADPH oxidase-derived superoxide might be a major factor in mediating the microglia-enhanced rotenone neurotoxicity. However, because of the relatively low specificity of these inhibitors, the exact source of superoxide induced by rotenone remains to be further determined. In this study, using primary mesencephalic cultures from NADPH oxidase--null (gp91phox-/-) or wild-type (gp91phox+/+) mice, we demonstrated a critical role for microglial NADPH oxidase in mediating microglia-enhanced rotenone neurotoxicity. In neuron--glia cultures, dopaminergic neurons from gp91phox-/- mice were more resistant to rotenone neurotoxicity than those from gp91phox+/+ mice. However, in neuron-enriched cultures, the neurotoxicity of rotenone was not different between the two types of mice. More importantly, the addition of microglia prepared from gp91phox+/+ mice but not from gp91phox-/- mice to neuron-enriched cultures markedly increased rotenone-induced degeneration of dopaminergic neurons. Furthermore, apocynin attenuated rotenone neurotoxicity only in the presence of microglia from gp91phox+/+ mice. These results indicated that the greatly enhanced neurotoxicity of rotenone was attributed to the release of NADPH oxidase-derived superoxide from activated microglia. This study also suggested that microglial NADPH oxidase may be a promising target for PD treatment.
Figures
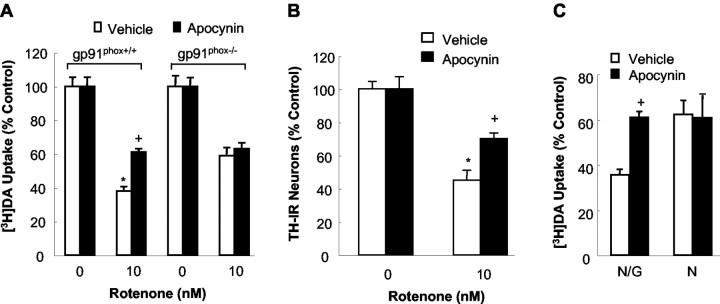
References
-
- Alam M, Schmidt WJ ( 2002) Rotenone destroys dopaminergic neurons and induces parkinsonian symptoms in rats. Behav Brain Res 136: 317—324. - PubMed
-
- Aloisi F ( 1999) The role of microglia and astrocytes in CNS immune surveillance and immunopathology. Adv Exp Med Biol 468: 123—133. - PubMed
-
- Babior BM ( 1999) NADPH oxidase: an update. Blood 93: 1464 —1476. - PubMed
-
- Betarbet R, Sherer TB, MacKenzie G, Garcia-Osuna M, Panov AV, Greenamyre JT ( 2000) Chronic systemic pesticide exposure reproduces features of Parkinson's disease. Nat Neurosci 3: 1301—1306. - PubMed
-
- Cassarino DS, Fall CP, Swerdlow RH, Smith TS, Halvorsen EM, Miller SW, Parks JP, Parker Jr WD, Bennett Jr JP ( 1997) Elevated reactive oxygen species and antioxidant enzyme activities in animal and cellular models of Parkinson's disease. Biochim Biophys Acta 1362: 77—86. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous