

Does cAMP response element-binding protein have a pivotal role in hippocampal synaptic plasticity and hippocampus-dependent memory?
- PMID: 12867515
- PMCID: PMC6740551
- DOI: 10.1523/JNEUROSCI.23-15-06304.2003
Does cAMP response element-binding protein have a pivotal role in hippocampal synaptic plasticity and hippocampus-dependent memory?
Abstract
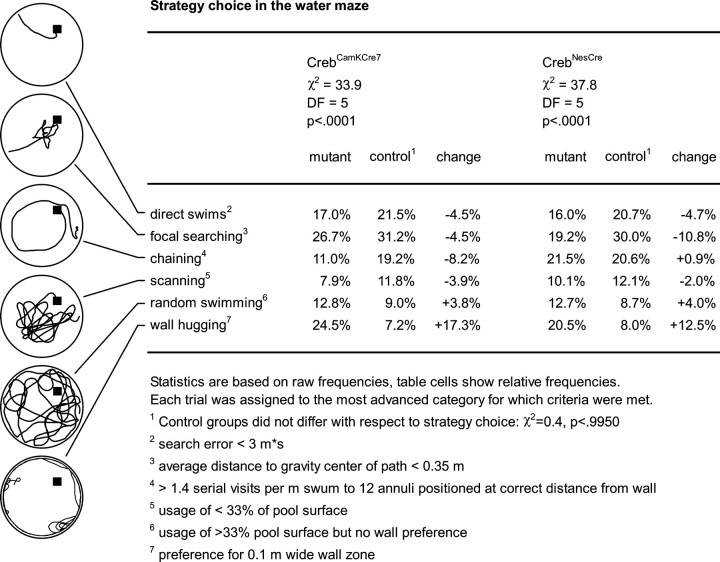
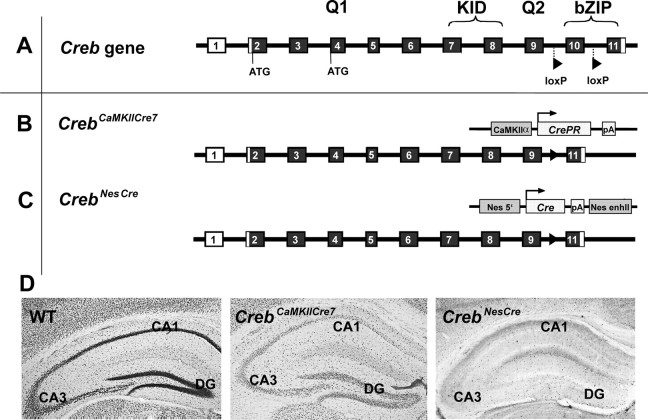
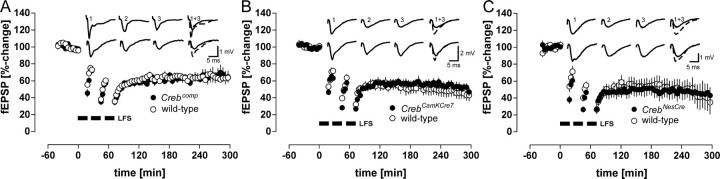
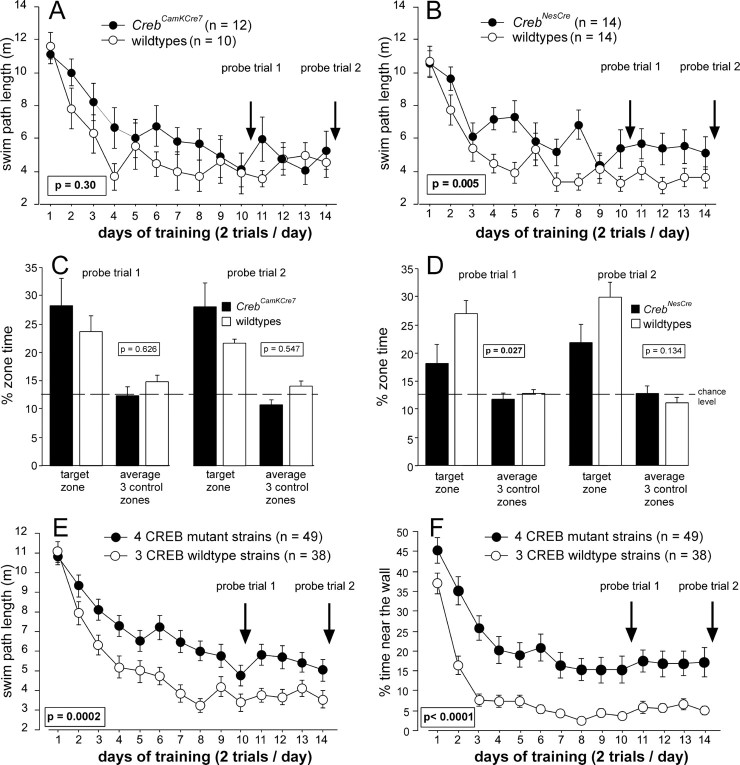
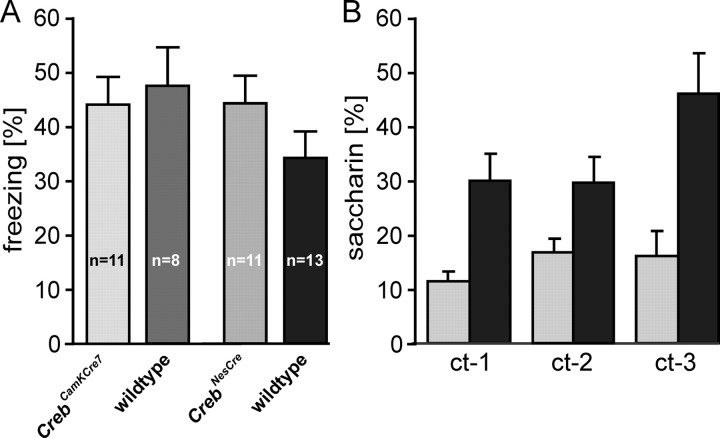
Previous studies addressing the role of the transcription factor cAMP response element-binding protein (CREB) in mammalian long-term synaptic plasticity and memory by gene targeting were compromised by incomplete deletion of the CREB isoforms. Therefore, we generated conditional knock-out strains with a marked reduction or complete deletion of all CREB isoforms in the hippocampus. In these strains, no deficits could be detected in lasting forms of hippocampal long-term potentiation (LTP) and long-term depression (LTD). When tested for hippocampus-dependent learning, mutants showed normal context-dependent fear conditioning. Water maze learning was impaired during the early stages, but many mutants showed satisfactory scores in probe trials thought to measure hippocampus-dependent spatial memory. However, conditioned taste aversion learning, a putatively hippocampus-independent memory test, was markedly impaired. Our data indicate that in the adult mouse brain, loss of CREB neither prevents learning nor substantially affects performance in some hippocampus-dependent tasks. Furthermore, it spares LTP and LTD in paradigms that are sensitive enough to detect deficits in other mutants. This implies either a species-specific or regionally restricted role of CREB in the brain and/or a compensatory upregulation of the cAMP response element modulator (CREM) and other as yet unidentified transcription factors.
Figures

References
-
- Agranoff BW, Davis RE, Brink JJ ( 1966) Chemical studies on memory fixation in goldfish. Brain Res 1: 303–309. - PubMed
-
- Ahn S, Ginty DD, Linden DJ ( 1999) A late phase of cerebellar long-term depression requires activation of CaMKIV and CREB. Neuron 23: 559–568. - PubMed
-
- Athos J, Impey S, Pineda VV, Chen X, Storm DR ( 2002) Hippocampal CRE-mediated gene expression is required for contextual memory formation. Nat Neurosci 5: 1119–1120. - PubMed
-
- Bartsch D, Ghirardi M, Skehel PA, Karl KA, Herder SP, Chen M, Bailey CH, Kandel ER ( 1995) Aplysia CREB2 represses long-term facilitation: relief of repression converts transient facilitation into long-term functional and structural change. Cell 83: 979–992. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases