

Immune complex-FcgammaR interaction modulates monocyte/macrophage molecules involved in inflammation and immune response
- PMID: 12869025
- PMCID: PMC1808765
- DOI: 10.1046/j.1365-2249.2003.02208.x
Immune complex-FcgammaR interaction modulates monocyte/macrophage molecules involved in inflammation and immune response
Abstract
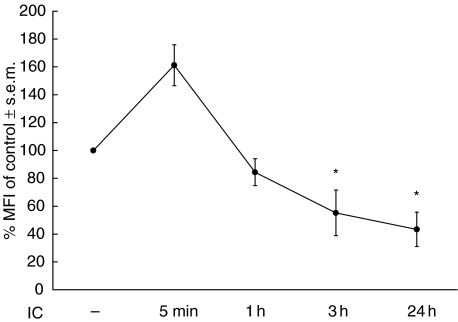
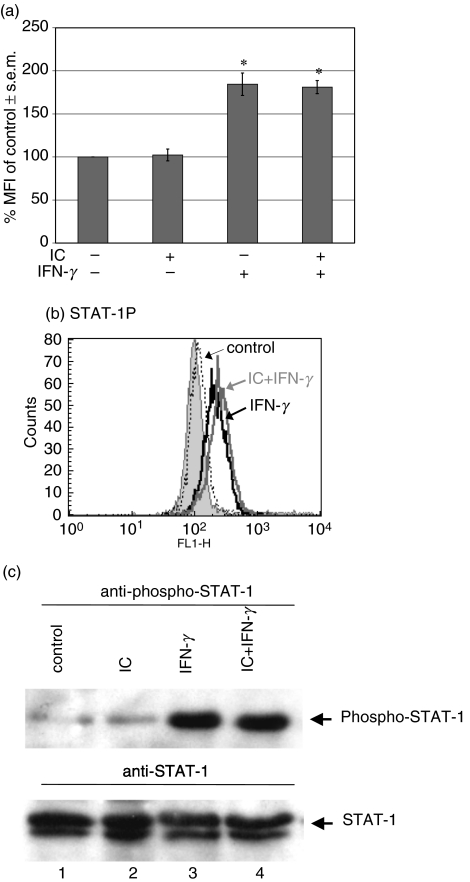
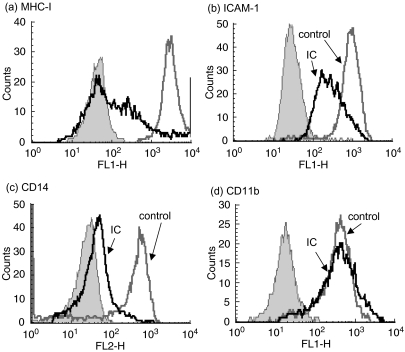
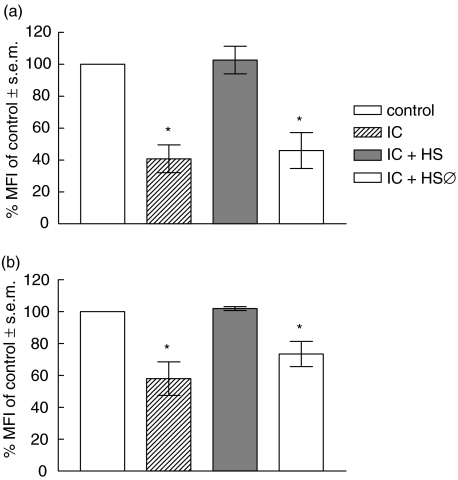
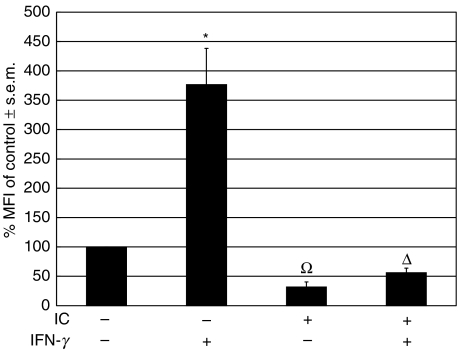
The interaction between receptors for the Fc portion of IgG (FcgammaRs) from monocytes/macrophages and immune complexes (IC) triggers regulatory and effector functions. Recently, we have demonstrated that IC exert a drastic inhibition of basal and IFN-gamma-induced expression of MHC class II on human monocytes. Taking into account that the regulation of MHC class II molecules is a crucial event in the immune response, in this report we extend our previous studies analysing the effect of STAT-1 phosphorylation in the down-regulatory process, the fate of the intracellular pool of MHC class II molecules and the effect of complement on MHC class II down-regulation induced by IC. We also studied the effect of IC on the expression of MHC class II (I-A(d)) in macrophages using a mouse model of chronic inflammation. We demonstrate that IC induce a depletion not only on surface expressed but also on intracellular MHC class II content and that IC-induced down-regulation of MHC class II is not mediated by the inhibition of STAT-1 phosphorylation. On the other hand, the effect of IC is not specific for the down-regulation of MHC class II, for it could be restricted to other molecules involved in inflammatory processes. Our experiments also show that the activation of the complement system could be a crucial step on the regulation of the effect of IC on MHC class II expression. In agreement with our in vitro experiments using human monocytes, IC treatment reduces the expression of MHC class II in a mouse model of chronic inflammation.
Figures
References
-
- Schifferli JA, Taylor RP. Physiological and pathological aspects of circulating immune complexes. Kidney Int. 1989;35:993–1003. - PubMed
-
- Mannik M. Pathophysiology of circulating immune complexes. Arthritis Rheumat. 1982;25:783–7. - PubMed
-
- Kasai S, Akaike T, Kunimoto T, Nitta K. Superoxide anion production from mouse peritoneal macrophages stimulated with surface-bound IgG and immune complexes: adsorption characteristics of IgG in relationship to biological activity. Eur J Immunol. 1982;12:1054–7. - PubMed
-
- Bonney RJ, Naruns P, Davies P, Humes JL. Antigen–antibody complexes stimulate the synthesis and release of prostaglandins by mouse peritoneal macrophages. Prostaglandins. 1979;18:605–16. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
