

Angiopoietin-related growth factor (AGF) promotes epidermal proliferation, remodeling, and regeneration
- PMID: 12871997
- PMCID: PMC170946
- DOI: 10.1073/pnas.1531901100
Angiopoietin-related growth factor (AGF) promotes epidermal proliferation, remodeling, and regeneration
Abstract
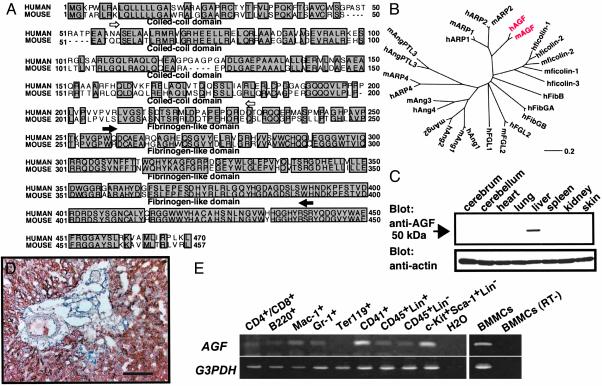
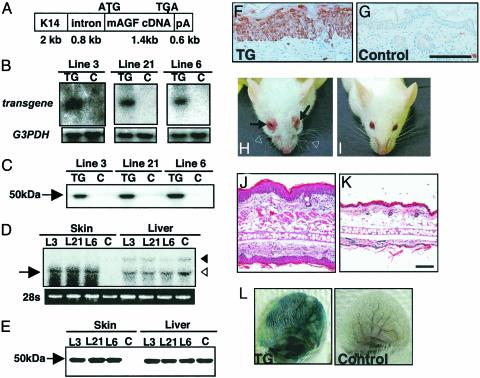
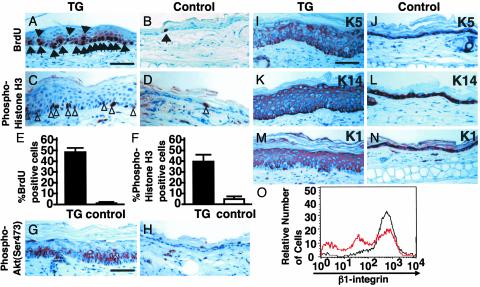
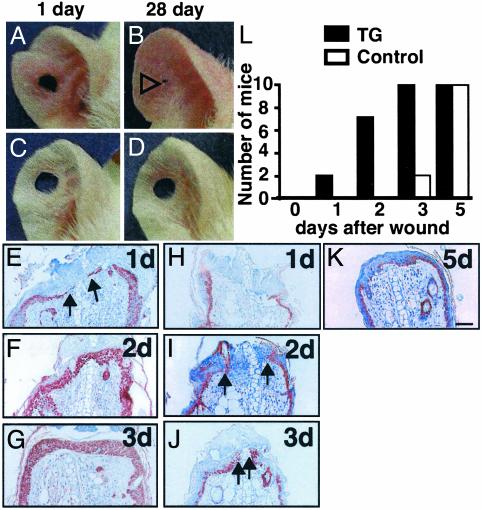
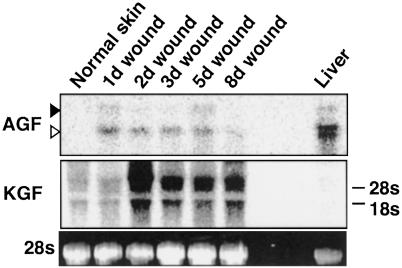
We report here the identification of an angiopoietin-related growth factor (AGF). To examine the biological function of AGF in vivo, we created transgenic mice expressing AGF in epidermal keratinocytes (K14-AGF). K14-AGF mice exhibited swollen and reddish ears, nose and eyelids. Histological analyses of K14-AGF mice revealed significantly thickened epidermis and a marked increase in proliferating epidermal cells as well as vascular cells in the skin compared with nontransgenic controls. In addition, we found rapid wound closure in the healing process and an unusual closure of holes punched in the ears of K14-AGF mice. Furthermore, we observed that AGF is expressed in platelets and mast cells, and detected at wounded skin, whereas there was no expression of AGF detected in normal skin tissues, suggesting that AGF derived from these infiltrated cells affects epidermal proliferation and thereby plays a role in the wound healing process. These findings demonstrate that biological functions of AGF in epidermal keratinocytes could lead to novel therapeutic strategies for wound care and epidermal regenerative medicine.
Figures
References
-
- Vassar, R. & Fuchs, E. (1991) Genes Dev. 5, 714-727. - PubMed
-
- Werner, S., Smola, H., Liao, X., Longaker, M. T., Krieg, T., Hofschneider, P. H. & Williams, L. T. (1994) Science 266, 819-822. - PubMed
-
- Mann, G. B., Fowler, K. J., Gabriel, A., Nice, E. C., Williams, R. L. & Dunn, A. R. (1993) Cell 73, 249-261. - PubMed
-
- Guo, L., Degensteim, L. & Fuchs, E. (1996) Genes Dev. 10, 165-175. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
