

Mutations in INVS encoding inversin cause nephronophthisis type 2, linking renal cystic disease to the function of primary cilia and left-right axis determination
- PMID: 12872123
- PMCID: PMC3732175
- DOI: 10.1038/ng1217
Mutations in INVS encoding inversin cause nephronophthisis type 2, linking renal cystic disease to the function of primary cilia and left-right axis determination
Abstract
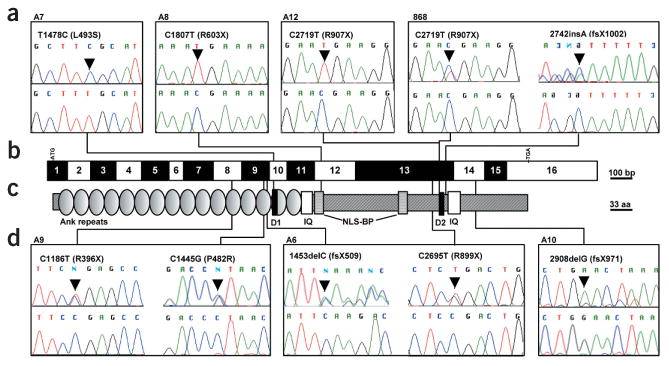
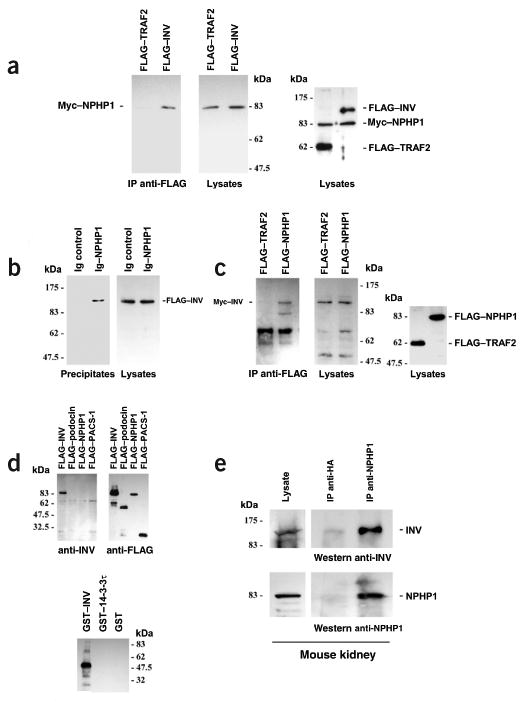
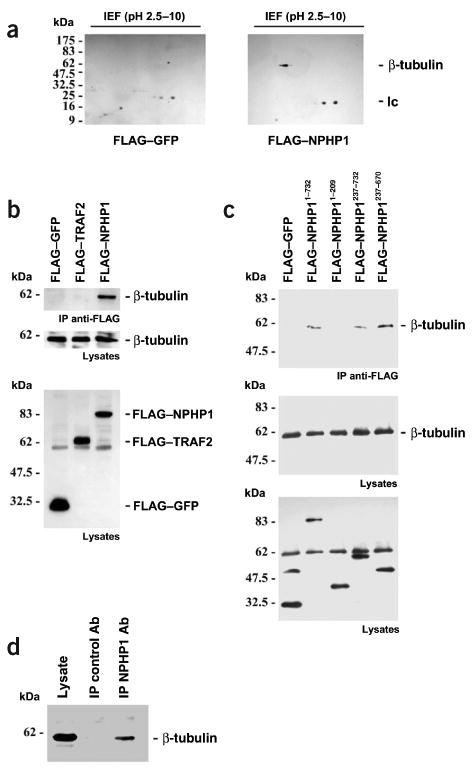
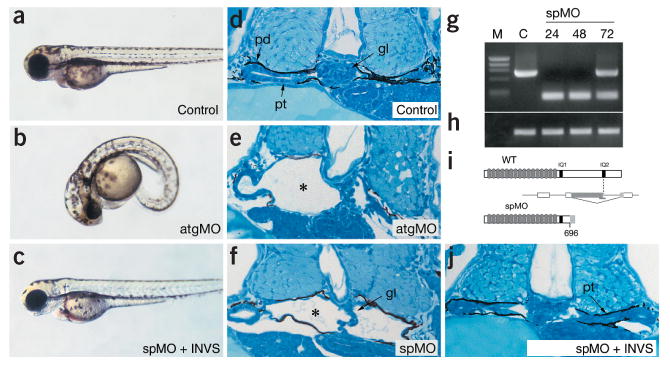
Nephronophthisis (NPHP), an autosomal recessive cystic kidney disease, leads to chronic renal failure in children. The genes mutated in NPHP1 and NPHP4 have been identified, and a gene locus associated with infantile nephronophthisis (NPHP2) was mapped. The kidney phenotype of NPHP2 combines clinical features of NPHP and polycystic kidney disease (PKD). Here, we identify inversin (INVS) as the gene mutated in NPHP2 with and without situs inversus. We show molecular interaction of inversin with nephrocystin, the product of the gene mutated in NPHP1 and interaction of nephrocystin with beta-tubulin, a main component of primary cilia. We show that nephrocystin, inversin and beta-tubulin colocalize to primary cilia of renal tubular cells. Furthermore, we produce a PKD-like renal cystic phenotype and randomization of heart looping by knockdown of invs expression in zebrafish. The interaction and colocalization in cilia of inversin, nephrocystin and beta-tubulin connect pathogenetic aspects of NPHP to PKD, to primary cilia function and to left-right axis determination.
Figures

Comment in
-
From cilia to cyst.Nat Genet. 2003 Aug;34(4):355-6. doi: 10.1038/ng0803-355. Nat Genet. 2003. PMID: 12923538 No abstract available.
References
-
- Smith C, Graham J. Congenital medullary cysts of the kidneys with severe refractory anemia. Am J Dis Child. 1945;69:369–377.
-
- Fanconi G, Hanhart E, Albertini A. Die familiäre juvenile Nephronophthise. Hel Pediatr Acta. 1951;6:1–49. - PubMed
-
- Hildebrandt F. Juvenile nephronophthisis. In: Barratt TM, Avner ED, Harmon WE, editors. Pediatric nephrology. Williams & Wilkins; Baltimore: 1999.
-
- Hildebrandt F, et al. A novel gene encoding an SH3 domain protein is mutated in nephronophthisis type 1. Nat Genet. 1997;17:149–153. - PubMed
-
- Saunier S, et al. A novel gene that encodes a protein with a putative src homology 3 domain is a candidate gene for familial juvenile nephronophthisis. Hum Mol Genet. 1997;6:2317–2323. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
