

Direct expansion of functional CD25+ CD4+ regulatory T cells by antigen-processing dendritic cells
- PMID: 12874257
- PMCID: PMC2194081
- DOI: 10.1084/jem.20030422
Direct expansion of functional CD25+ CD4+ regulatory T cells by antigen-processing dendritic cells
Abstract
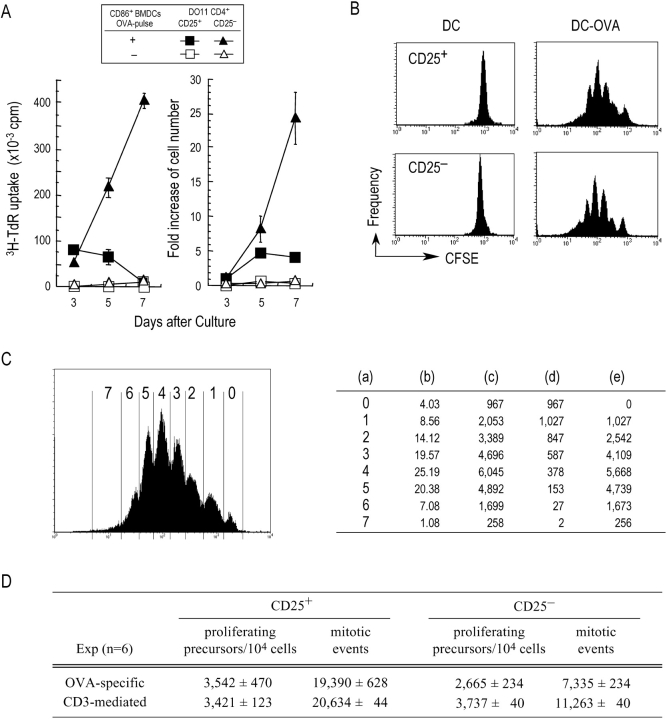
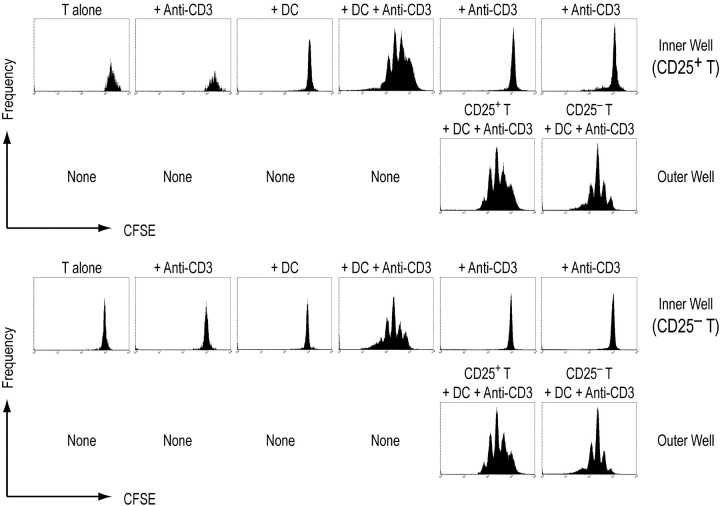
An important pathway for immune tolerance is provided by thymic-derived CD25+ CD4+ T cells that suppress other CD25- autoimmune disease-inducing T cells. The antigen-presenting cell (APC) requirements for the control of CD25+ CD4+ suppressor T cells remain to be identified, hampering their study in experimental and clinical situations. CD25+ CD4+ T cells are classically anergic, unable to proliferate in response to mitogenic antibodies to the T cell receptor complex. We now find that CD25+ CD4+ T cells can proliferate in the absence of added cytokines in culture and in vivo when stimulated by antigen-loaded dendritic cells (DCs), especially mature DCs. With high doses of DCs in culture, CD25+ CD4+ and CD25- CD4+ populations initially proliferate to a comparable extent. With current methods, one third of the antigen-reactive T cell receptor transgenic T cells enter into cycle for an average of three divisions in 3 d. The expansion of CD25+ CD4+ T cells stops by day 5, in the absence or presence of exogenous interleukin (IL)-2, whereas CD25- CD4+ T cells continue to grow. CD25+ CD4+ T cell growth requires DC-T cell contact and is partially dependent upon the production of small amounts of IL-2 by the T cells and B7 costimulation by the DCs. After antigen-specific expansion, the CD25+ CD4+ T cells retain their known surface features and actively suppress CD25- CD4+ T cell proliferation to splenic APCs. DCs also can expand CD25+ CD4+ T cells in the absence of specific antigen but in the presence of exogenous IL-2. In vivo, both steady state and mature antigen-processing DCs induce proliferation of adoptively transferred CD25+ CD4+ T cells. The capacity to expand CD25+ CD4+ T cells provides DCs with an additional mechanism to regulate autoimmunity and other immune responses.
Figures






References
-
- Sakaguchi, S., N. Sakaguchi, J. Shimizu, S. Yamazaki, T. Sakihama, M. Itoh, Y. Kuniyasu, T. Nomura, M. Toda, and T. Takahashi. 2001. Immunologic tolerance maintained by CD25+ CD4+ regulatory T cells: their common role in controlling autoimmunity, tumor immunity, and transplantation tolerance. Immunol. Rev. 182:18–32. - PubMed
-
- Shevach, E.M. 2000. Regulatory T cells in autoimmunity. Annu. Rev. Immunol. 18:423–449. - PubMed
-
- Saoudi, A., B. Seddon, V. Heath, V. Fowell, and D. Mason. 1996. The physiological role of regulatory T cells in the prevention of autoimmunity: the function of the thymus in the generation of the regulatory T cell subset. Immunol. Rev. 149:195–216. - PubMed
-
- Sakaguchi, S., N. Sakaguchi, M. Asano, M. Itoh, and M. Toda. 1995. Immunologic self-tolerance maintained by activated T cells expressing IL-2 receptor alpha-chains (CD25). Breakdown of a single mechanism of self-tolerance causes various autoimmune diseases. J. Immunol. 155:1151–1164. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
