

Dendritic cells are responsible for the capacity of CpG oligodeoxynucleotides to act as an adjuvant for protective vaccine immunity against Leishmania major in mice
- PMID: 12874261
- PMCID: PMC2194077
- DOI: 10.1084/jem.20030645
Dendritic cells are responsible for the capacity of CpG oligodeoxynucleotides to act as an adjuvant for protective vaccine immunity against Leishmania major in mice
Abstract
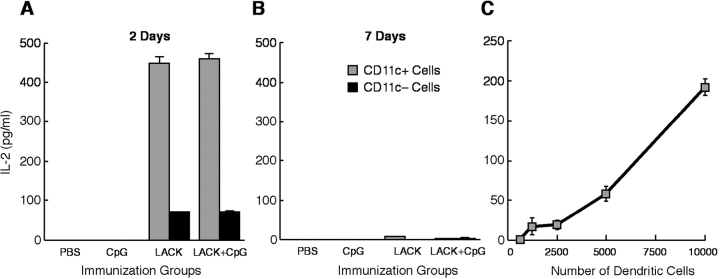
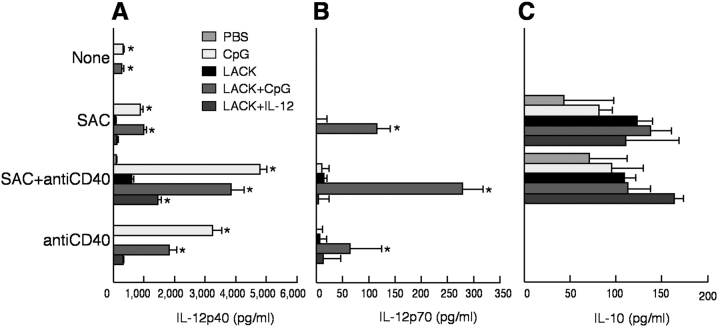
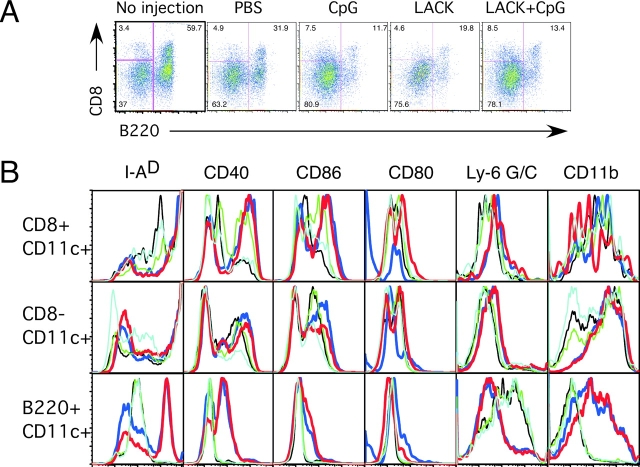
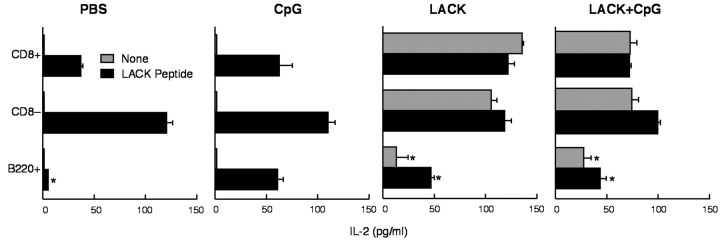
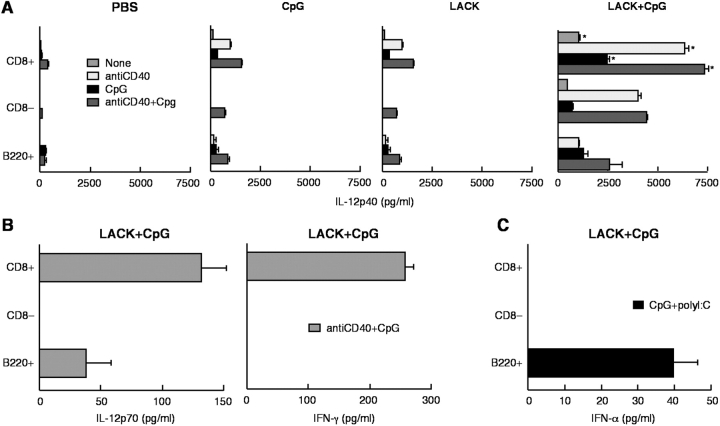
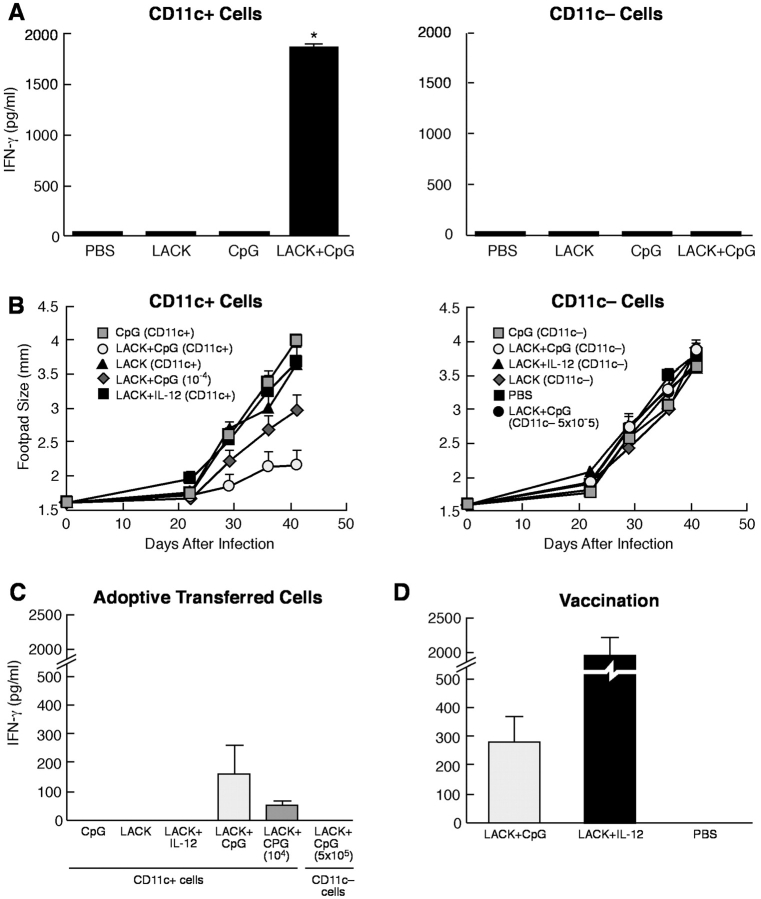
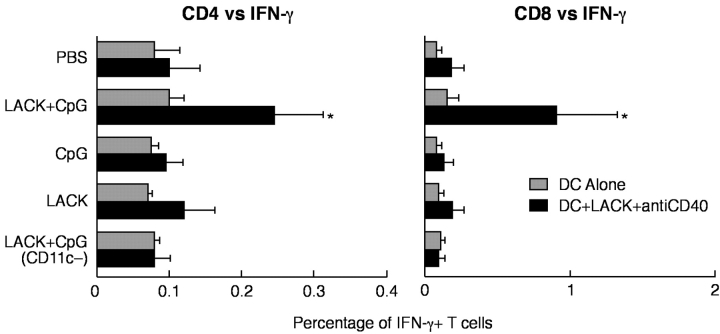
Vaccination with leishmanial Ag and CpG oligodeoxynucleotides (ODN) confers sustained cellular immunity and protection to infectious challenge up to 6 mo after immunization. To define the cellular mechanism by which CpG ODN mediate their adjuvant effects in vivo, the functional capacity of distinct dendritic cell (DC) subsets was assessed in the lymph nodes (LNs) of BALB/c mice, 36 h after immunization with the leishmanial antigen (LACK) and CpG ODN. After this immunization, there was a striking decrease in the frequency of the CD11c+B220+ plasmacytoid DCs with a proportionate increase in CD11c+CD8-B220- cells. CD11c+CD8+B220- cells were the most potent producers of interleukin (IL)-12 p70 and interferon (IFN)-gamma, while plasmacytoid DCs were the only subset capable of secreting IFN-alpha. In terms of antigen presenting capacity, plasmacytoid DCs were far less efficient compared with the other DC subsets. To certify that DCs were responsible for effective vaccination, we isolated CD11c+ and CD11c- cells 36 h after immunization and used such cells to elicit protective immunity after adoptive transfer in naive, Leishmania major susceptible BALB/c mice. CD11c+ cells but not 10-fold higher numbers of CD11c- cells from such immunized mice mediated protection. Therefore, the combination of LACK antigen and CpG ODN adjuvant leads to the presence of CD11c+ DCs in the draining LN that are capable of vaccinating naive mice in the absence of further antigen or adjuvant.
Figures
References
-
- Seder, R.A., and A.V. Hill. 2000. Vaccines against intracellular infections requiring cellular immunity. Nature. 406:793–798. - PubMed
-
- O'Garra, A. 1998. Cytokines induce the development of functionally heterogeneous T helper cell subsets. Immunity. 8:275–283. - PubMed
-
- Murphy, K.M., W. Ouyang, J.D. Farrar, J. Yang, S. Ranganath, H. Asnagli, M. Afkarian, and T.L. Murphy. 2000. Signaling and transcription in T helper development. Annu. Rev. Immunol. 18:451–494. - PubMed
-
- Krieg, A.M. 2002. CpG motifs in bacterial DNA and their immune effects. Annu. Rev. Immunol. 20:709–760. - PubMed
-
- Barton, G.M., and R. Medzhitov. 2002. Control of adaptive immune responses by Toll-like receptors. Curr. Opin. Immunol. 14:380–383. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
