

Low dielectric permittivity of water at the membrane interface: effect on the energy coupling mechanism in biological membranes
- PMID: 12885673
- PMCID: PMC1303247
- DOI: 10.1016/S0006-3495(03)74565-2
Low dielectric permittivity of water at the membrane interface: effect on the energy coupling mechanism in biological membranes
Erratum in
- Biophys J. 2003 Sep;85(3):2069
Abstract
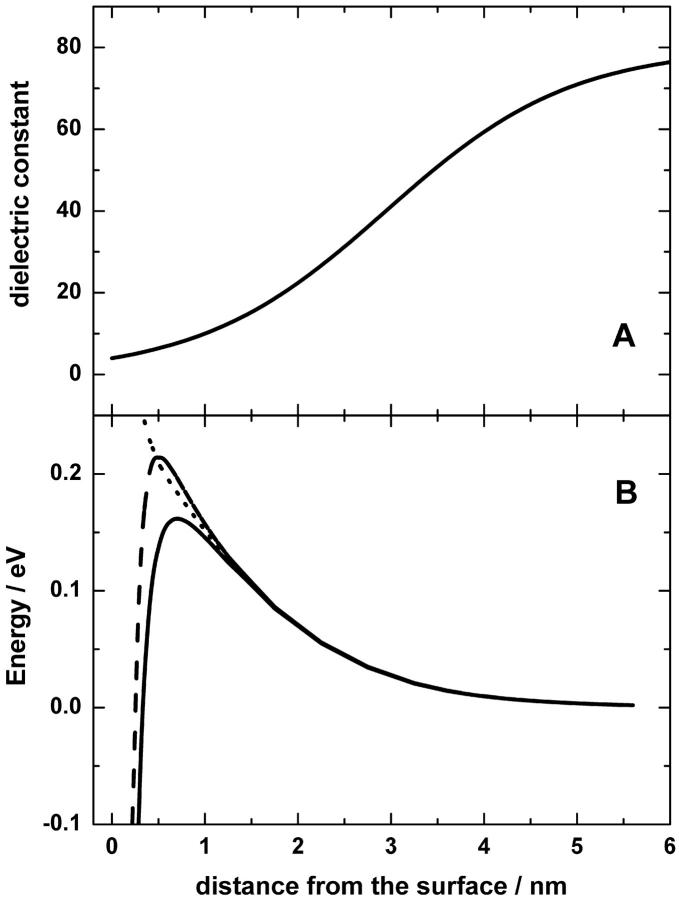
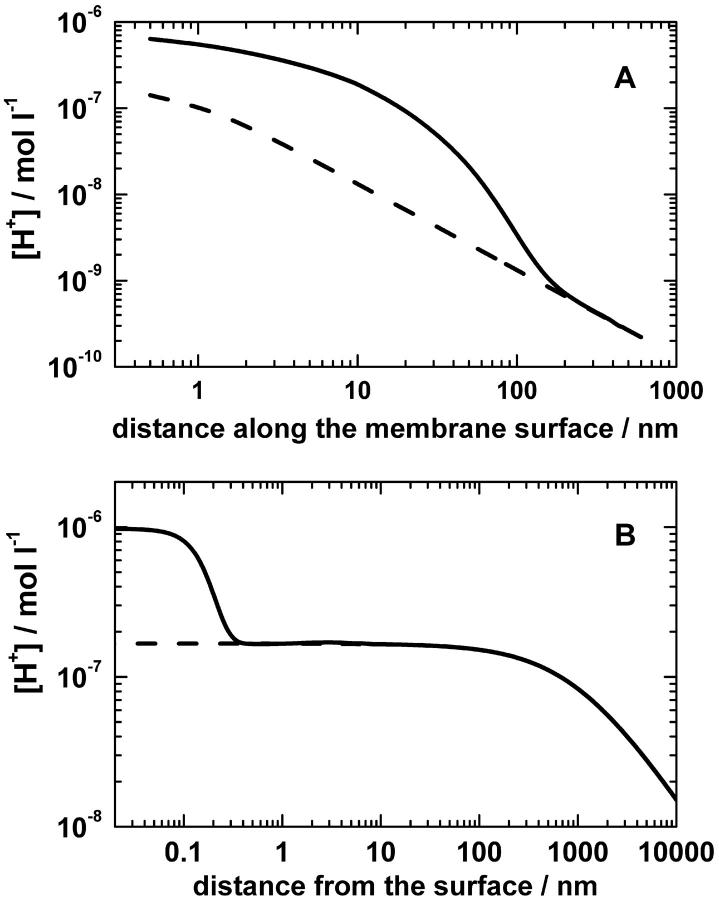
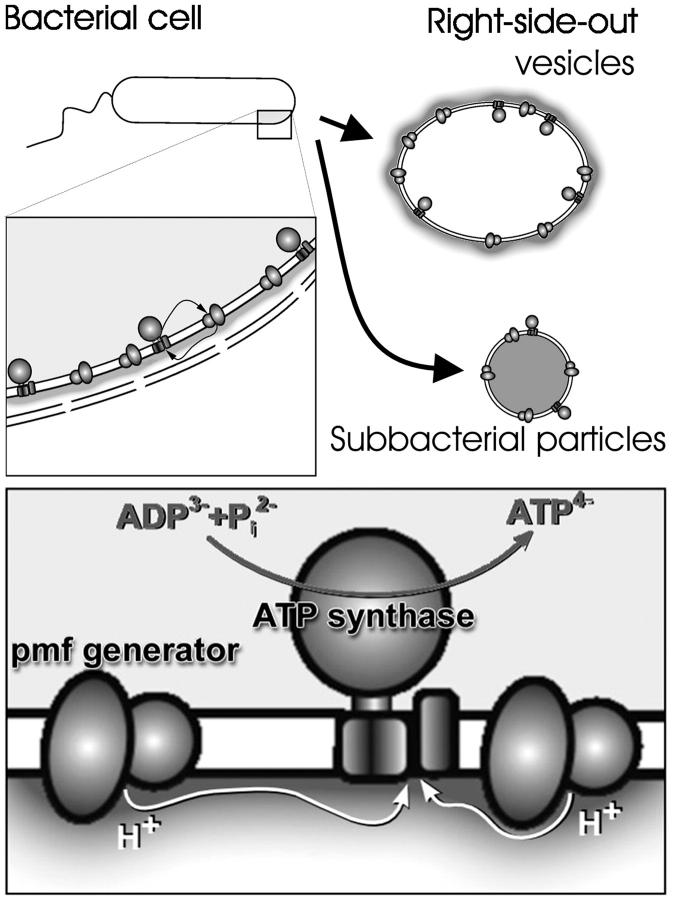
Protonmotive force (the transmembrane difference in electrochemical potential of protons, ) drives ATP synthesis in bacteria, mitochondria, and chloroplasts. It has remained unsettled whether the entropic (chemical) component of relates to the difference in the proton activity between two bulk water phases (deltapH(B)) or between two membrane surfaces (deltapH(S)). To scrutinize whether deltapH(S) can deviate from deltapH(B), we modeled the behavior of protons at the membrane/water interface. We made use of the surprisingly low dielectric permittivity of interfacial water as determined by O. Teschke, G. Ceotto, and E. F. de Souza (O. Teschke, G. Ceotto, and E. F. de Sousa, 2001, PHYS: Rev. E. 64:011605). Electrostatic calculations revealed a potential barrier in the water phase some 0.5-1 nm away from the membrane surface. The barrier was higher for monovalent anions moving toward the surface (0.2-0.3 eV) than for monovalent cations (0.1-0.15 eV). By solving the Smoluchowski equation for protons spreading away from proton "pumps" at the surface, we found that the barrier could cause an elevation of the proton concentration at the interface. Taking typical values for the density of proton pumps and for their turnover rate, we calculated that a potential barrier of 0.12 eV yielded a steady-state pH(S) of approximately 6.0; the value of pH(S) was independent of pH in the bulk water phase under neutral and alkaline conditions. These results provide a rationale to solve the long-lasting problem of the seemingly insufficient protonmotive force in mesophilic and alkaliphilic bacteria.
Figures
References
-
- Adams, J. 1993. MUDPACK-2: multigrid software for elliptic partial differential equations on uniform grids with any resolution. Appl. Math. Comp. 53:235–249.
-
- Antonenko, Y. N., O. N. Kovbasnjuk, and L. S. Yaguzhinsky. 1993. Evidence in favor of the existence of a kinetic barrier for proton transfer from a surface of bilayer phospholipid membrane to bulk water. Biochim. Biophys. Acta. 1150:45–50. - PubMed
-
- Arata, H., I. Takenaka, and M. Nishimura. 1987. Flash-induced proton release in Rhodopseudomonas sphaeroides spaeroplasts. J. Biochem. 101:261–265. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
