

Functional characterization of cis- and trans-regulatory elements involved in expression of phospholipid hydroperoxide glutathione peroxidase
- PMID: 12888488
- PMCID: PMC169948
- DOI: 10.1093/nar/gkg650
Functional characterization of cis- and trans-regulatory elements involved in expression of phospholipid hydroperoxide glutathione peroxidase
Abstract
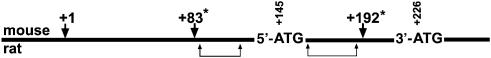
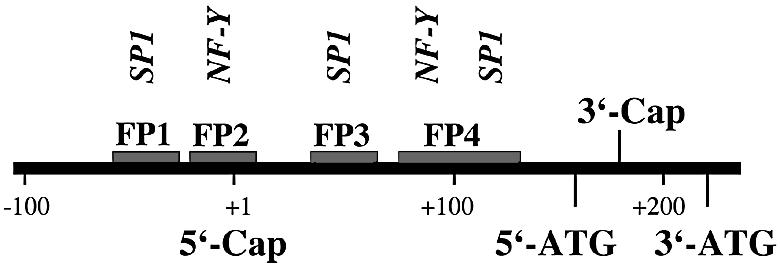
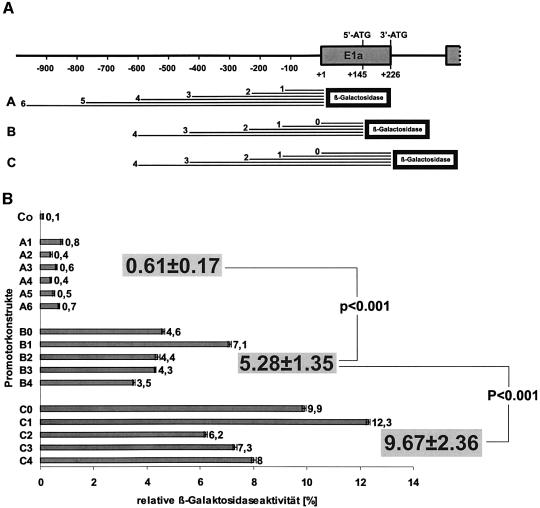
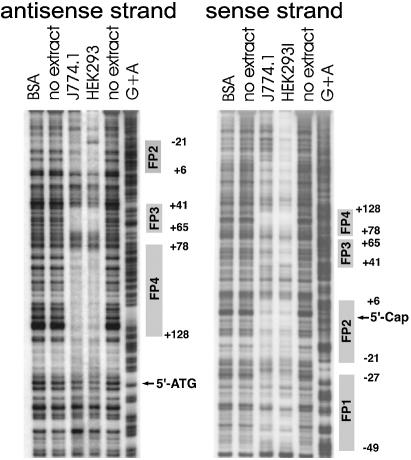
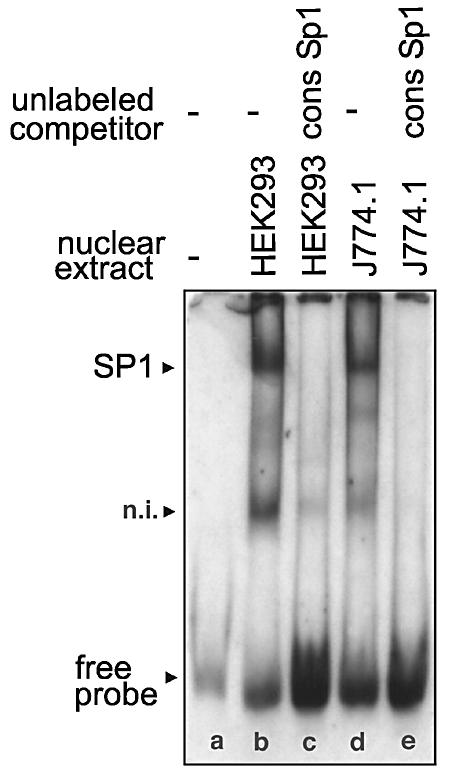
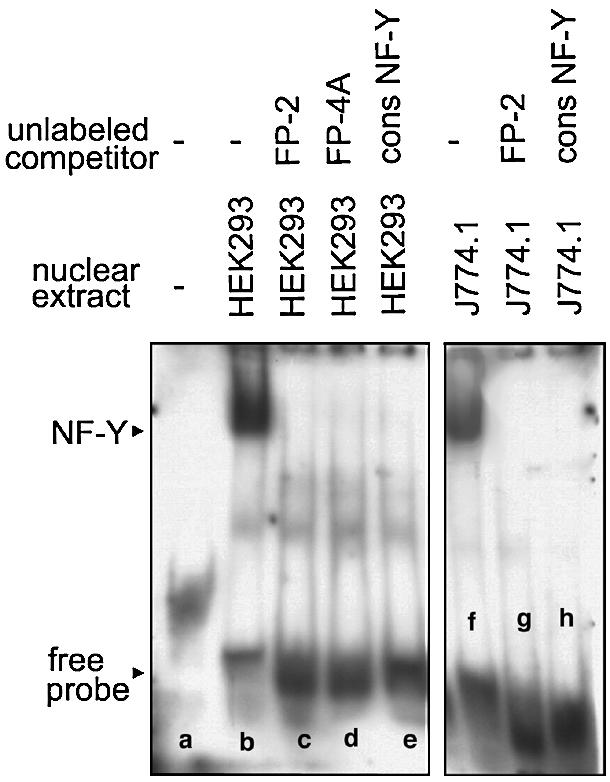
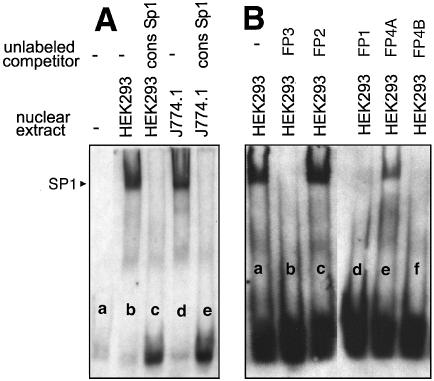
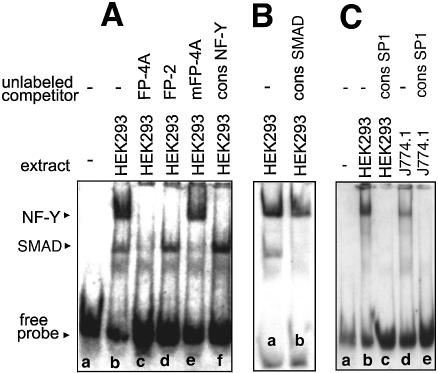
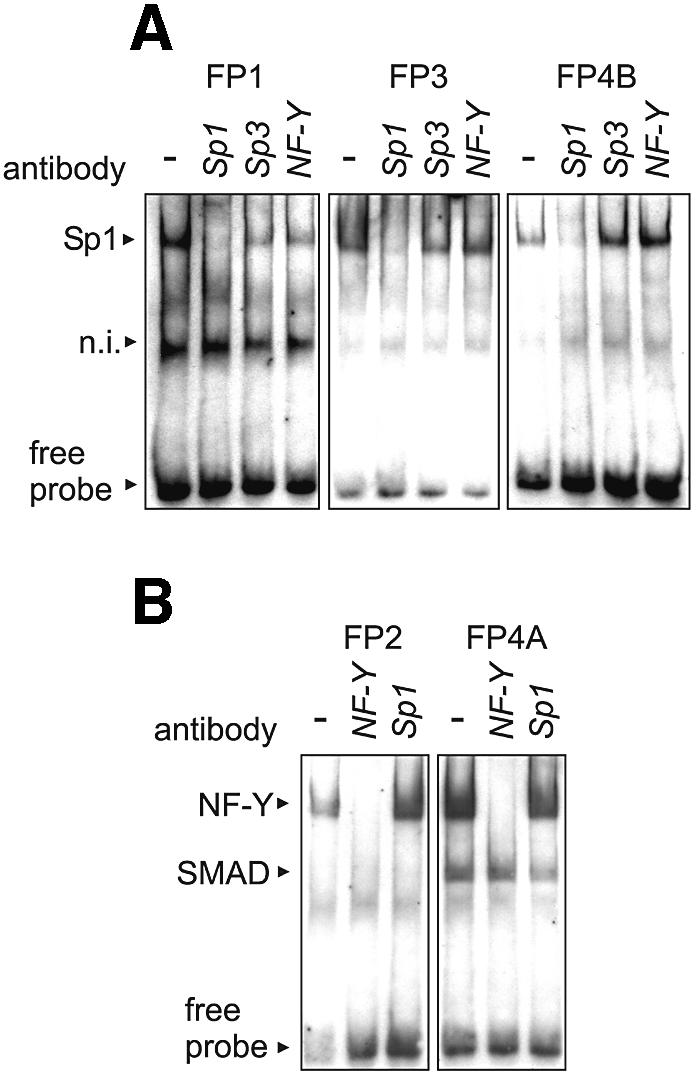
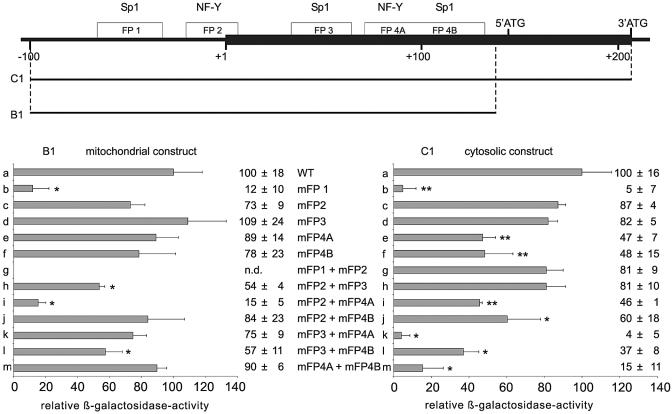
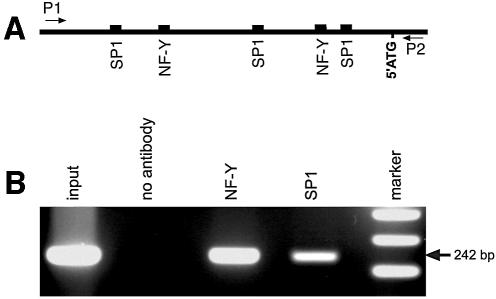
Phospholipid hydroperoxide glutathione peroxidase (phGPx) is a member of the seleno glutathione peroxidase family that is comprised of five selenoproteins capable of reducing hydroperoxy lipids to the corresponding alcohols. The enzyme has been implicated in antioxidative defense, but its high expression level in testicular tissue suggests a more specific function during sperm maturation. The phGPx is encoded for by a joint sperm nucleus/phGPx gene (sn/phGPx) and can be expressed as a mitochondrial or cytosolic isoform. Although sn/phGPx genes have been cloned from various mammalian species expression regulation of the enzyme has not been studied in detail. We investigated the 5'-flanking region of the murine sn/phGPx gene and observed basic promoter activity in a 200 bp region localized immediately upstream of the translational initiation site of the cytosolic isoform (3'-ATG). DNase protection assays indicated the presence of five distinct protein-binding regions and electrophoretic mobility shift assays and supershift experiments revealed binding of stimulating protein 1 (SP1), nuclear factor Y (NF-Y) and members of the SMAD family. Site-directed mutagenesis of the consensus binding sequences abolished in vitro transcription factor binding. Expression of reporter genes was most effectively impaired when SP1/SP3 and NF-Y binding site-deficient constructs were tested. Chromatin immunoprecipitation suggested the in vivo relevance of these transcription factors. Our data indicate that the basic phGPx promoter constitutes a 200 bp oligonucleotide, which is localized immediately upstream of the 3'-ATG and involves functional SP1/SP3, NF-Y and SMAD binding sites. The corresponding trans-regulatory proteins may contribute to differential expression regulation of the mitochondrial and cytosolic phGPx isoforms.
Figures
Similar articles
-
The CCAAT-box binding factor NF-Y is required for the expression of phospholipid hydroperoxide glutathione peroxidase in human epidermoid carcinoma A431 cells.FEBS Lett. 1999 Jul 16;455(1-2):111-6. doi: 10.1016/s0014-5793(99)00866-2. FEBS Lett. 1999. PMID: 10428483
-
Identification of the positive regulatory and distinct core regions of promoters, and transcriptional regulation in three types of mouse phospholipid hydroperoxide glutathione peroxidase.J Biochem. 2006 Oct;140(4):573-90. doi: 10.1093/jb/mvj186. Epub 2006 Sep 7. J Biochem. 2006. PMID: 16959796
-
Regulation of expression of the phospholipid hydroperoxide/sperm nucleus glutathione peroxidase gene. Tissue-specific expression pattern and identification of functional cis- and trans-regulatory elements.J Biol Chem. 2003 Jan 24;278(4):2571-80. doi: 10.1074/jbc.M209064200. Epub 2002 Nov 8. J Biol Chem. 2003. PMID: 12427732
-
[Biological significance of lipid hydroperoxide and its reducing enzyme, phospholipid hydroperoxide glutathione peroxidase, in mammalian cells].Yakugaku Zasshi. 2004 Dec;124(12):937-57. doi: 10.1248/yakushi.124.937. Yakugaku Zasshi. 2004. PMID: 15577264 Review. Japanese.
-
Role of mitochondrial phospholipid hydroperoxide glutathione peroxidase (PHGPx) as an antiapoptotic factor.Biol Pharm Bull. 2004 Jul;27(7):956-60. doi: 10.1248/bpb.27.956. Biol Pharm Bull. 2004. PMID: 15256721 Review.
Cited by
-
Reduced expression of the ferroptosis inhibitor glutathione peroxidase-4 in multiple sclerosis and experimental autoimmune encephalomyelitis.J Neurochem. 2019 Feb;148(3):426-439. doi: 10.1111/jnc.14604. Epub 2018 Dec 3. J Neurochem. 2019. PMID: 30289974 Free PMC article.
-
GPX3 expression was down-regulated but positively correlated with poor outcome in human cancers.Front Oncol. 2023 Feb 9;13:990551. doi: 10.3389/fonc.2023.990551. eCollection 2023. Front Oncol. 2023. PMID: 36845676 Free PMC article.
-
A DNA replication-related element downstream from the initiation site of Drosophila selenophosphate synthetase 2 gene is essential for its transcription.Nucleic Acids Res. 2004 Apr 30;32(8):2482-93. doi: 10.1093/nar/gkh569. Print 2004. Nucleic Acids Res. 2004. PMID: 15121905 Free PMC article.
-
The GI-GPx gene is a target for Nrf2.Mol Cell Biol. 2005 Jun;25(12):4914-23. doi: 10.1128/MCB.25.12.4914-4923.2005. Mol Cell Biol. 2005. PMID: 15923610 Free PMC article.
-
Loss of the SIN3 transcriptional corepressor results in aberrant mitochondrial function.BMC Biochem. 2010 Jul 9;11:26. doi: 10.1186/1471-2091-11-26. BMC Biochem. 2010. PMID: 20618965 Free PMC article.
References
-
- Flohé L. (1989) The selenoprotein glutathione peroxidase. In Dolphin,D., Poulson,R. and Avramovic,O. (eds), Glutathione: Chemical, Biochemical and Medical Aspects—Part A. John Wiley and Sons, New York, pp. 643–731.
-
- Behne D. and KyriakopoulosA. (2001) Mammalian selenium-containing proteins. Annu. Rev. Nutr., 21, 453–473. - PubMed
-
- Kuhn H. and Borchert,A. (2002) Regulation of enzymatic lipid peroxidation. The interplay of peroxidizing and peroxide reducing enzymes. Free Radic. Biol. Med., 33, 154–172. - PubMed
-
- Ursini F., Maiorino,M., Valente,M., Ferri,L. and Gregolin,C. (1982) Purification from pig liver of a protein which protects liposomes and biomembranes from peroxidative degradation and exhibits glutathione peroxidase activity on phosphatidylcholine hydroperoxides. Biochim. Biophys. Acta, 710, 197–211. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
