

Lack of huntingtin-associated protein-1 causes neuronal death resembling hypothalamic degeneration in Huntington's disease
- PMID: 12890790
- PMCID: PMC6740731
- DOI: 10.1523/JNEUROSCI.23-17-06956.2003
Lack of huntingtin-associated protein-1 causes neuronal death resembling hypothalamic degeneration in Huntington's disease
Abstract
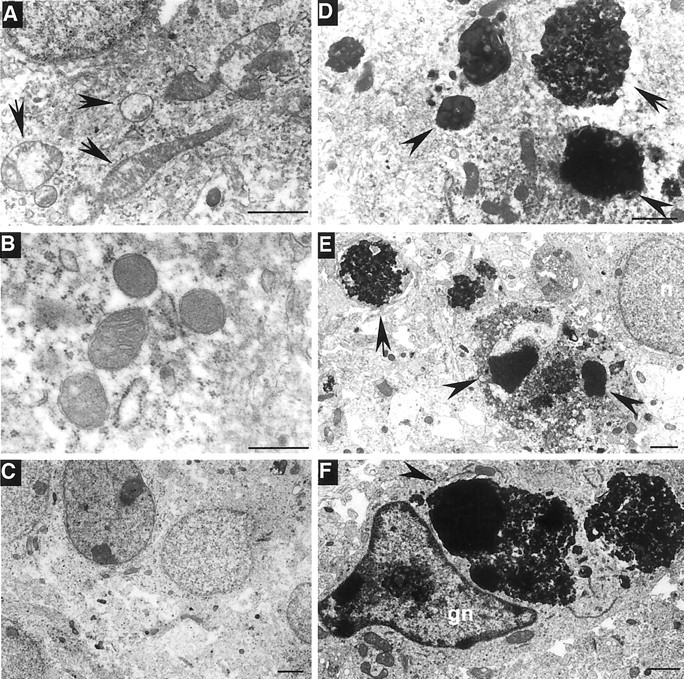
Huntington's disease (HD) is caused by a polyglutamine expansion in the disease protein huntingtin. The polyglutamine expansion causes huntingtin to interact abnormally with a number of proteins. However, it is unclear whether, and how, huntingtin-associated proteins are involved in the neurodegeneration in HD. Here, we show that huntingtin-associated protein-1 (HAP1), which is involved in intracellular trafficking of epidermal growth factor receptor (EGFR), is highly expressed in the hypothalamus. Mice lacking HAP1 die after birth because of depressed feeding activity. Terminal deoxynucleotidyl transferase-mediated biotinylated UTP nick end labeling staining and electron microscopic examination revealed the degeneration in hypothalamic regions that control feeding behavior. Hypothalamic degeneration was also observed in HD transgenic mice that have a significant loss of body weight. Inhibition of HAP1 expression decreases EGFR signaling and cell viability, whereas overexpression of HAP1 enhances this signaling activity and inhibits mutant huntingtin-mediated cytotoxicity. These results suggest that the effect of mutant huntingtin on HAP1 and EGFR signaling may contribute to the hypothalamic neurodegeneration and loss of body weight in HD.
Figures






Similar articles
-
Huntingtin-associated protein-1 deficiency in orexin-producing neurons impairs neuronal process extension and leads to abnormal behavior in mice.J Biol Chem. 2010 May 21;285(21):15941-9. doi: 10.1074/jbc.M110.107318. Epub 2010 Mar 19. J Biol Chem. 2010. PMID: 20304926 Free PMC article.
-
Mutant huntingtin causes context-dependent neurodegeneration in mice with Huntington's disease.J Neurosci. 2003 Mar 15;23(6):2193-202. doi: 10.1523/JNEUROSCI.23-06-02193.2003. J Neurosci. 2003. PMID: 12657678 Free PMC article.
-
Mutant huntingtin and glycogen synthase kinase 3-beta accumulate in neuronal lipid rafts of a presymptomatic knock-in mouse model of Huntington's disease.J Neurosci Res. 2010 Jan;88(1):179-90. doi: 10.1002/jnr.22184. J Neurosci Res. 2010. PMID: 19642201
-
[Huntington's disease: intracellular signaling pathways and neuronal death].J Soc Biol. 2005;199(3):247-51. doi: 10.1051/jbio:2005026. J Soc Biol. 2005. PMID: 16471265 Review. French.
-
Mechanisms of neuronal cell death in Huntington's disease.Cytogenet Genome Res. 2003;100(1-4):287-95. doi: 10.1159/000072864. Cytogenet Genome Res. 2003. PMID: 14526190 Review.
Cited by
-
The Role of Hypothalamic Pathology for Non-Motor Features of Huntington's Disease.J Huntingtons Dis. 2019;8(4):375-391. doi: 10.3233/JHD-190372. J Huntingtons Dis. 2019. PMID: 31594240 Free PMC article. Review.
-
Huntingtin-associated protein-1 deficiency in orexin-producing neurons impairs neuronal process extension and leads to abnormal behavior in mice.J Biol Chem. 2010 May 21;285(21):15941-9. doi: 10.1074/jbc.M110.107318. Epub 2010 Mar 19. J Biol Chem. 2010. PMID: 20304926 Free PMC article.
-
Huntingtin-associated protein 1 regulates exocytosis, vesicle docking, readily releasable pool size and fusion pore stability in mouse chromaffin cells.J Physiol. 2014 Apr 1;592(7):1505-18. doi: 10.1113/jphysiol.2013.268342. Epub 2013 Dec 23. J Physiol. 2014. PMID: 24366265 Free PMC article.
-
Huntingtin associated protein 1 and its functions.Cell Adh Migr. 2009 Jan-Mar;3(1):71-6. doi: 10.4161/cam.3.1.7511. Epub 2009 Jan 26. Cell Adh Migr. 2009. PMID: 19262167 Free PMC article. Review.
-
The ubiquitin proteasome system in glia and its role in neurodegenerative diseases.Front Mol Neurosci. 2014 Aug 8;7:73. doi: 10.3389/fnmol.2014.00073. eCollection 2014. Front Mol Neurosci. 2014. PMID: 25152710 Free PMC article. Review.
References
-
- Block-Galarza J, Chase KO, Sapp E, Vaughn KT, Vallee RB, DiFiglia M, Aronin N ( 1997) Fast transport and retrograde movement of huntingtin and HAP 1 in axons. NeuroReport 8: 2247-2251. - PubMed
-
- Cattaneo E, Rigamonti D, Goffredo D, Zuccato C, Squitieri F, Sipione S ( 2001) Loss of normal huntingtin function: new developments in Huntington's disease research. Trends Neurosci 24: 182-188. - PubMed
-
- Chan EY, Nasir J, Gutekunst CA, Coleman S, Maclean A, Maas A, Metzler M, Gertsenstein M, Ross CA, Nagy A, Hayden MR ( 2002) Targeted disruption of huntingtin-associated protein-1 (Hap1) results in postnatal death due to depressed feeding behavior. Hum Mol Genet 11: 945-959. - PubMed
-
- Chen RH, Chang MC, Su YH, Tsai YT, Kuo ML ( 1999) Interleukin-6 inhibits transforming growth factor-beta-induced apoptosis through the phosphatidylinositol 3-kinase/Akt and signal transducers and activators of transcription 3 pathways. J Biol Chem 274: 23013-23019. - PubMed
-
- Davies SW, Turmaine M, Cozens BA, DiFiglia M, Sharp AH, Ross CA, Scherzinger E, Wanker EE, Mangiarini L, Bates GP ( 1997) Formation of neuronal intranuclear inclusions underlies the neurological dysfunction in mice transgenic for the HD mutation. Cell 90: 537-548. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous