

Dependence of Helicobacter pylori urease activity on the nickel-sequestering ability of the UreE accessory protein
- PMID: 12896998
- PMCID: PMC166491
- DOI: 10.1128/JB.185.16.4787-4795.2003
Dependence of Helicobacter pylori urease activity on the nickel-sequestering ability of the UreE accessory protein
Abstract
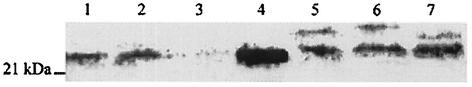
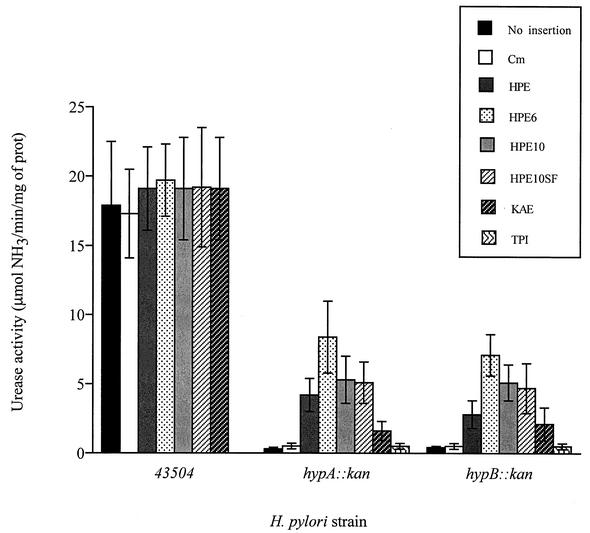
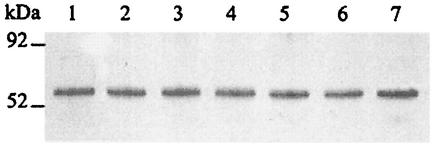
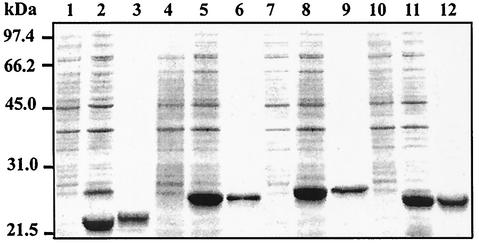
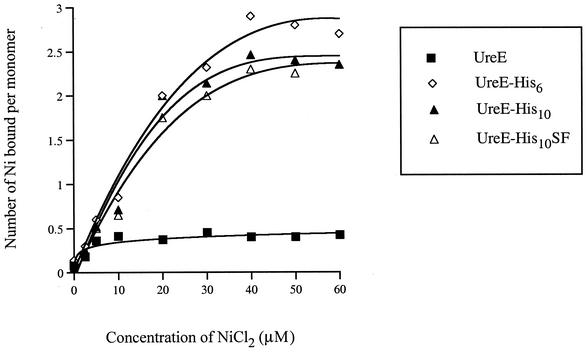
The Helicobacter pylori ureE gene product was previously shown to be required for urease expression, but its characteristics and role have not been determined. The UreE protein has now been overexpressed in Escherichia coli, purified, and characterized, and three altered versions were expressed to address a nickel-sequestering role of UreE. Purified UreE formed a dimer in solution and was capable of binding one nickel ion per dimer. Introduction of an extra copy of ureE into the chromosome of mutants carrying mutations in the Ni maturation proteins HypA and HypB resulted in partial restoration of urease activity (up to 24% of the wild-type levels). Fusion proteins of UreE with increased ability to bind nickel were constructed by adding histidine-rich sequences (His-6 or His-10 to the C terminus and His-10 as a sandwich fusion) to the UreE protein. Each fusion protein was overexpressed in E. coli and purified, and its nickel-binding capacity and affinity were determined. Each construct was also expressed in wild-type H. pylori and in hypA and hypB mutant strains for determining in vivo urease activities. The urease activity was increased by introduction of all the engineered versions, with the greatest Ni-sequestering version (the His-6 version) also conferring the greatest urease activity on both the hypA and hypB mutants. The differences in urease activities were not due to differences in the amounts of urease peptides. Addition of His-6 to another expressed protein (triose phosphate isomerase) did not result in stimulation of urease, so urease activation is not related to the level of nonspecific protein-bound nickel. The results indicate a correlation between H. pylori urease activity and the nickel-sequestering ability of the UreE accessory protein.
Figures

References
-
- Akada, J. K., M. Shirai, H. Takeuchi, M. Tsuda, and T. Nakazawa. 2000. Identification of the urease operon in Helicobacter pylori and its control by mRNA decay in response to pH. Mol. Microbiol. 36:1071-1084. - PubMed
-
- Blaser, M. J. 1990. Helicobacter pylori and the pathogenesis of gastroduodenal inflammation. J. Infect. Dis. 161:626-633. - PubMed
-
- Ciurli, S., N. Safarov, S. Miletti, A. Dikiy, S. K. Christensen, K. Kornetzky, D. A. Bryant, I. Vandenberghe, B. Devreese, B. Samyn, H. Remaut, and J. van Beeumen. 2002. Molecular characterization of Bacillus pasteurii UreE, a metal-binding chaperone for the assembly of the urease active site. J. Biol. Inorg. Chem. 7:623-631. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
