

A conserved structural motif reveals the essential transcriptional repression function of Spen proteins and their role in developmental signaling
- PMID: 12897056
- PMCID: PMC196244
- DOI: 10.1101/gad.266203
A conserved structural motif reveals the essential transcriptional repression function of Spen proteins and their role in developmental signaling
Abstract
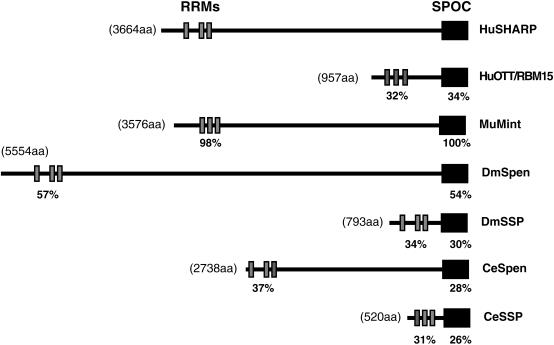
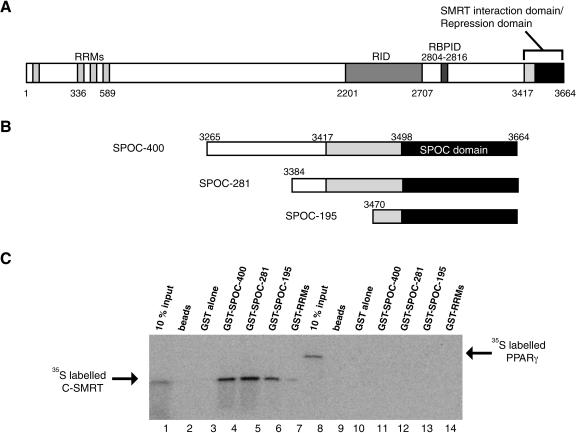
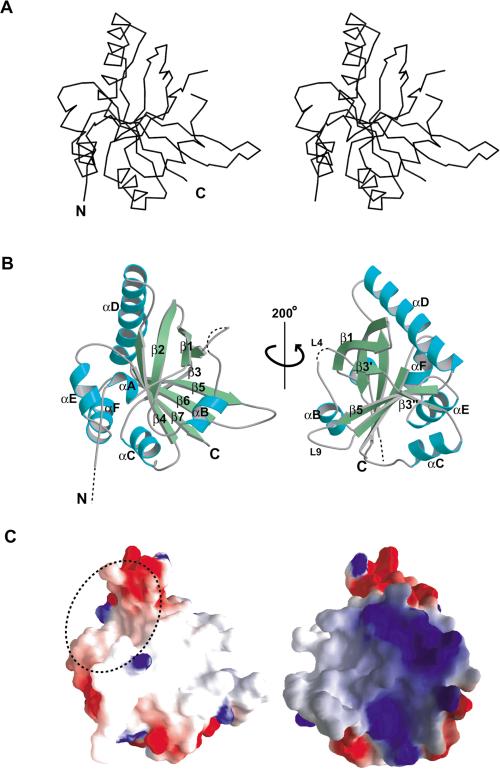
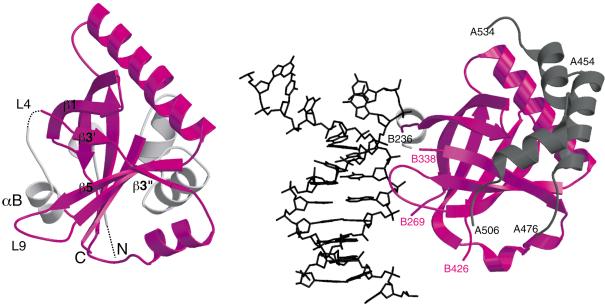
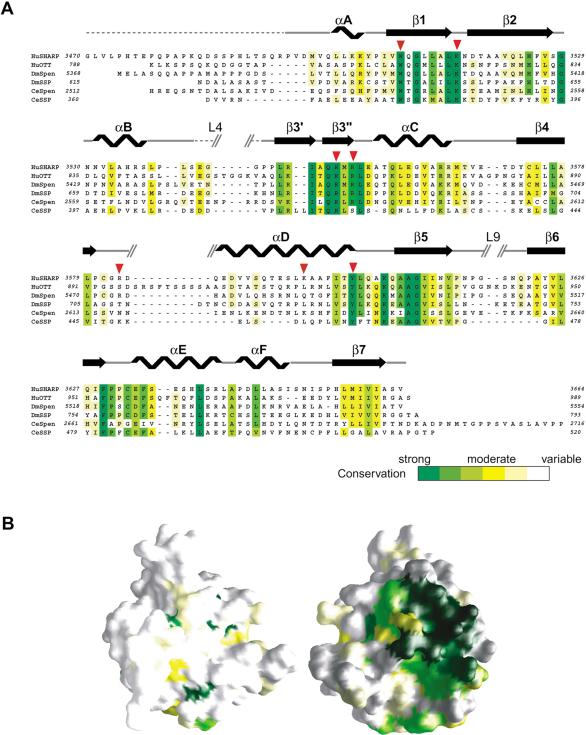
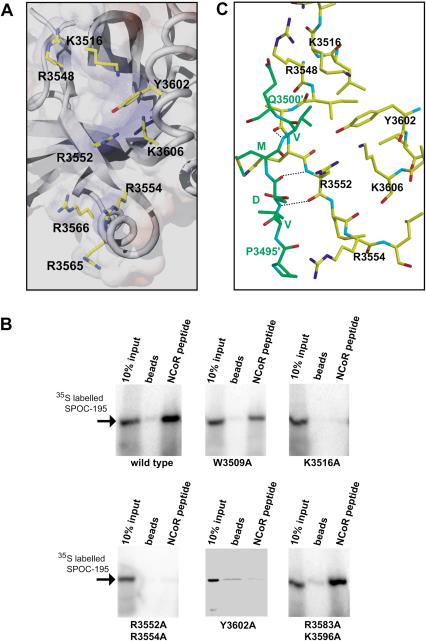
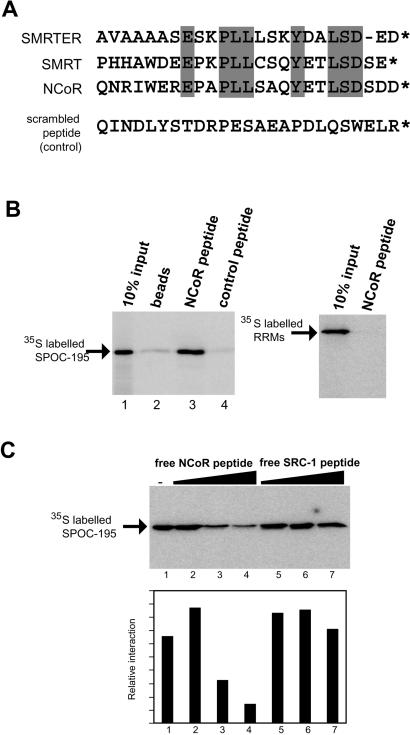
Spen proteins regulate the expression of key transcriptional effectors in diverse signaling pathways. They are large proteins characterized by N-terminal RNA-binding motifs and a highly conserved C-terminal SPOC domain. The specific biological role of the SPOC domain (Spen paralog and ortholog C-terminal domain), and hence, the common function of Spen proteins, has been unclear to date. The Spen protein, SHARP (SMRT/HDAC1-associated repressor protein), was identified as a component of transcriptional repression complexes in both nuclear receptor and Notch/RBP-Jkappa signaling pathways. We have determined the 1.8 A crystal structure of the SPOC domain from SHARP. This structure shows that essentially all of the conserved surface residues map to a positively charged patch. Structure-based mutational analysis indicates that this conserved region is responsible for the interaction between SHARP and the universal transcriptional corepressor SMRT/NCoR (silencing mediator for retinoid and thyroid receptors/nuclear receptor corepressor. We demonstrate that this interaction involves a highly conserved acidic motif at the C terminus of SMRT/NCoR. These findings suggest that the conserved function of the SPOC domain is to mediate interaction with SMRT/NCoR corepressors, and that Spen proteins play an essential role in the repression complex.
Figures
References
-
- Bertrand N., Castro, D.S., and Guillemot, F. 2002. Proneural genes and the specification of neural cell types. Nat. Rev. Neurosci. 3: 517-530. - PubMed
-
- Brunger A.T., Adams, P.D., Clore, G.M., DeLano, W.L., Gros, P., Grosse-Kunstleve, R.W., Jiang, J.S., Kuszewski, J., Nilges, M., Pannu, N.S., et al. 1998. Crystallography & NMR system: A new software suite for macromolecular structure determination. Acta Crystallogr. D. Biol. Crystallogr. 54: 905-921. - PubMed
-
- Chen F. and Rebay, I. 2000. split ends, a new component of the Drosophila EGF receptor pathway, regulates development of midline glial cells. Curr. Biol. 10: 943-946. - PubMed
-
- Chen J.D. and Evans, R.M. 1995. A transcriptional co-repressor that interacts with nuclear hormone receptors. Nature 377: 454-457. - PubMed
-
- Collaborative Computational Project, Number 4. 1994. The CCP4 suite: Programs for protein crystallography. Acta Crystallogr. D. Biol. Crystallogr. 50: 760-763. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous