

doi: 10.1128/MCB.23.16.5606-5613.2003.
Stem-loop IV of tetrahymena telomerase RNA stimulates processivity in trans
Affiliations
- PMID: 12897134
- PMCID: PMC166324
- DOI: 10.1128/MCB.23.16.5606-5613.2003
Item in Clipboard
Stem-loop IV of tetrahymena telomerase RNA stimulates processivity in trans
Mol Cell Biol.
2003 Aug.
Abstract
Telomerase is a ribonucleoprotein enzyme responsible for the addition of telomeres onto the ends of chromosomes. Short or dysfunctional telomeres can lead to cell growth arrest, apoptosis, and genomic instability. Telomerase uses its RNA subunit to copy a short template region for telomere synthesis. To probe for regions of Tetrahymena telomerase RNA essential for function, we assayed 27 circularly permuted RNA deletions for telomerase in vitro activity and binding to the telomerase reverse transcriptase catalytic protein subunit. We found that stem-loop IV is required for wild-type telomerase activity in vitro and will stimulate processivity when added in trans.
Figures
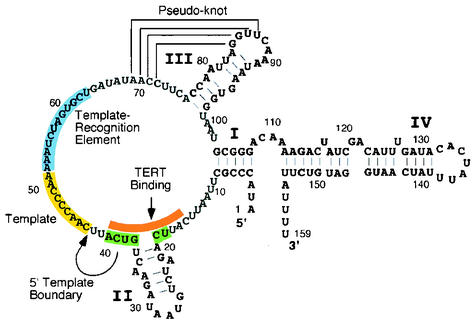
Summary of the functional regions identified in Tetrahymena telomerase RNA. The Tetrahymena RNA contains four helical regions, stem I to IV, and a conserved pseudoknot structure (brackets) (30). The specific helical regions are marked with a Roman numeral, and the nucleotide positions for each region are marked every 10 bases beginning at the 5′end. The template region is highlighted in yellow. A sequence conserved in ciliates that defines the template boundary is highlighted in green (1). This sequence overlaps with the defined tTERT-binding region shown in orange (19, 20). A region important for template utilization, the template-recognition element, is highlighted in blue (28).
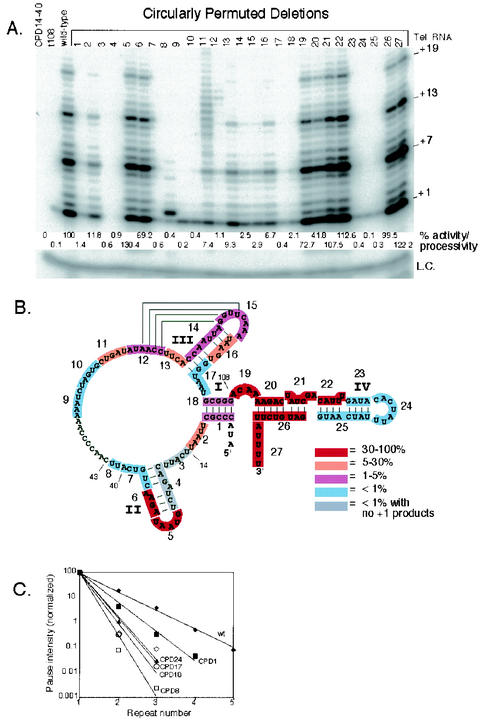
Telomerase activity reconstituted with telomerase RNA CPDs. (A) Telomerase activity assays were reconstituted with the telomerase RNAs indicated in the figure. +1, +7, +13, and +19 on the right indicate telomerase extension products from the d(GGGGTT)3 primer. Quantified telomerase activity/processivity is shown below each lane expressed as % of wild-type (see methods). L.C., loading control added for all reactions. (B) Summary of the reconstituted telomerase activity levels of all 27 CPDs shown with the telomerase RNA secondary structure. The highlighted sequence indicates the RNA region deleted in each CPD. The CPD number is shown next to each highlighted RNA sequence from 5′ to 3′ along the RNA. The color indicates the relative level of telomerase activity. Red (50 to 100%) indicates wild-type activity. Orange (5 to 50%), purple (1 to 5%), and light blue (less than 1%) indicate activity less than that of the wild type. Gray indicates no activity, since no products at +1 were observed. Relevant telomerase RNA nucleotide positions are indicated on the figure. (C) Telomerase processivity quantification of selected CPDs. The intensity of each major band (+1, +7, +13, +19, and +25) from telomerase assay in panel A was quantified as described in the Materials and Methods and plotted for each repeat number. The processivity of each mutant is inversely related to the slope of the line.
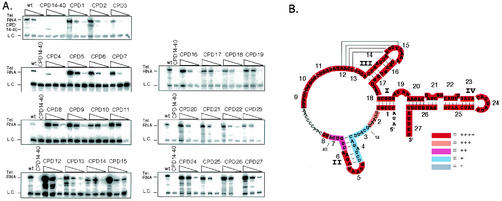
tTERT binding of telomerase RNA circularly permuted deletions (CPDs). (A) Wild-type RNA and CPDs tested for tTERT binding are indicated in the figure. Note the CPD14-40 RNA migrates significantly faster than wild-type and CPD RNAs due to the larger deletion of residues. For each set of binding reactions three different RNA concentrations were used (100, 10, and 1 mM from left to right). Except for the top panel (CPDs 1 to 3), wild-type and CPD14-40 telomerase RNAs were only tested at the 10 mM RNA concentration as positive and negative controls. (B) Summary of the tTERT binding to telomerase RNA CPDs compared to wild-type shown with the telomerase RNA secondary structure. The highlighted sequence indicates the RNA region deleted in each CPD. The CPD number is shown next to each highlighted RNA sequence from 5′ to 3′ along the RNA. The color indicates the relative level of tTERT binding. Red (++++) indicates wild-type binding. Orange (+++), purple (++), and light blue (+) indicate binding less than that of wild-type tTERT. Gray indicates no tTERT binding. Relevant telomerase RNA nucleotide positions are indicated on the figure.
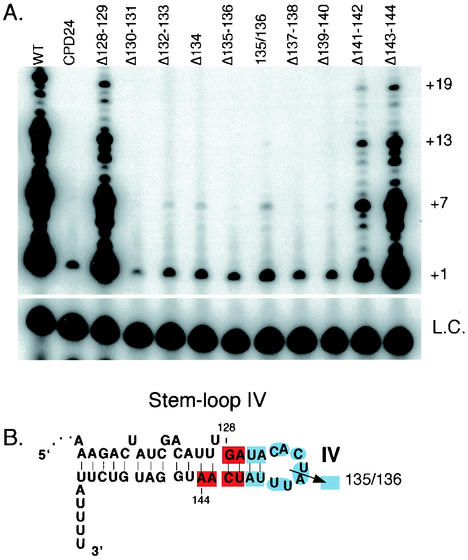
Telomerase activity reconstituted with small telomerase RNA circularly permuted deletions (CPDs) within stem-loop IV. (A) Telomerase activity assays were reconstituted with wild-type or mutant telomerase RNAs as indicated in the figure. +1, +7, +13, and +19 on the right indicate telomerase extension products from the d(GGGGTT)3 primer. L.C., loading control added to each reaction. (B) Summary of the reconstituted telomerase activity levels for the stem-loop IV CPDs. The residue numbers 128 and 144 indicate the RNA region targeted for smaller deletions. Each highlighted sequence indicates the RNA residues deleted in each smaller CPD. The color indicates the relative level of reconstituted activity; red indicates wild-type or near-wild-type activity, and light blue indicates low and less processive activity, similar to that of CPD 24. The arrow between residues 135 and 136 indicates relative level of reconstituted activity for a circular permutation that breaks the phosphodiester bond between residues 135 and 136 (135/136).
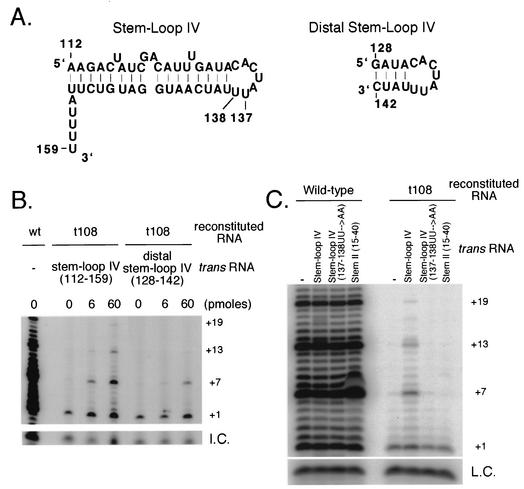
Stem-loop IV stimulates telomerase processivity. (A) Secondary structures of stem-loop IV nt 112 to 159 (left) and distal stem-loop IV nt 128 to 142 (right) are shown. Residue numbers are indicated on the structures. (B) Telomerase reconstituted with wild-type (wt) or t108 telomerase RNA was assayed for telomerase activity in the presence of 0, 6, or 60 pmol of stem-loop IV RNA (nt 112 to 159) or distal stem-loop IV RNA (nt 128 to 142). +1, +7, +13, and +19 on the right indicate telomerase extension products from the d(GGGGTT)3 primer. I.C., internal control for loading. (C) Telomerase reconstituted with wild-type or t108 telomerase RNA was assayed for telomerase activity alone or in the presence of 500 pmol of stem-loop IV, stem-loop IV (137-138 UU→AA) double point mutant or stem II nt 15 to 40. +1, +7, +13, and +19 on the right indicate telomerase extension products from the d(GGGGTT)3 primer. L.C., loading control for telomerase reactions.
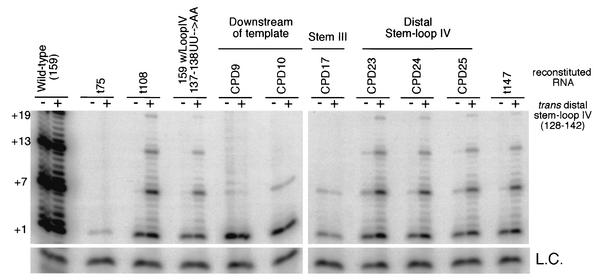
Stem-loop IV in trans rescues a specific subset of CPDs with low processivity. (A) Telomerase was reconstituted with various telomerase RNA CPDs and mutants as indicated in the figure. +1, +7,+13, and +19 on the left indicate telomerase extension products from the d(GGGGTT)3 primer. For telomerase CPDs, the corresponding RNA region is indicated above each deletion or group of deletions. Each reconstituted telomerase was assayed for telomerase activity in the absence or presence of 500 pmol of distal stem-loop IV (nt 128 to 142). L.C., loading control for telomerase reactions.
Similar articles
-
Roles for RNA in telomerase nucleotide and repeat addition processivity.Mol Cell. 2003 Jun;11(6):1673-83. doi: 10.1016/s1097-2765(03)00232-6. Mol Cell. 2003. PMID: 12820978 Free PMC article.
-
Structure of stem-loop IV of Tetrahymena telomerase RNA.EMBO J. 2006 Jul 12;25(13):3156-66. doi: 10.1038/sj.emboj.7601195. Epub 2006 Jun 15. EMBO J. 2006. PMID: 16778765 Free PMC article.
-
Template boundary definition in Tetrahymena telomerase.Genes Dev. 2002 Feb 15;16(4):415-20. doi: 10.1101/gad.962602. Genes Dev. 2002. PMID: 11850404 Free PMC article.
-
Structure and function of telomerase RNA.Curr Opin Struct Biol. 2006 Jun;16(3):307-18. doi: 10.1016/j.sbi.2006.05.005. Epub 2006 May 18. Curr Opin Struct Biol. 2006. PMID: 16713250 Review.
-
Telomerase Mechanism of Telomere Synthesis.Annu Rev Biochem. 2017 Jun 20;86:439-460. doi: 10.1146/annurev-biochem-061516-045019. Epub 2017 Jan 30. Annu Rev Biochem. 2017. PMID: 28141967 Free PMC article. Review.
Cited by
-
Structural study of elements of Tetrahymena telomerase RNA stem-loop IV domain important for function.RNA. 2006 Aug;12(8):1475-85. doi: 10.1261/rna.112306. Epub 2006 Jun 29. RNA. 2006. PMID: 16809815 Free PMC article.
-
The structure of an enzyme-activating fragment of human telomerase RNA.RNA. 2005 Apr;11(4):394-403. doi: 10.1261/rna.7222505. Epub 2005 Feb 9. RNA. 2005. PMID: 15703438 Free PMC article.
-
Characterisation of the Arabidopsis thaliana telomerase TERT-TR complex.Plant Mol Biol. 2024 May 14;114(3):56. doi: 10.1007/s11103-024-01461-w. Plant Mol Biol. 2024. PMID: 38743198 Free PMC article.
-
The RNA accordion model for template positioning by telomerase RNA during telomeric DNA synthesis.Nat Struct Mol Biol. 2011 Nov 20;18(12):1371-5. doi: 10.1038/nsmb.2174. Nat Struct Mol Biol. 2011. PMID: 22101935 Free PMC article.
-
Telomerase: an RNP enzyme synthesizes DNA.Cold Spring Harb Perspect Biol. 2011 May 1;3(5):a003558. doi: 10.1101/cshperspect.a003558. Cold Spring Harb Perspect Biol. 2011. PMID: 20660025 Free PMC article. Review.
References
-
- Autexier, C., and C. W. Greider. 1995. Boundary elements of the Tetrahymena telomerase RNA template and alignment domains. Genes Dev. 9:2227-2239. - PubMed
-
- Beattie, T. L., W. Zhou, M. O. Robinson, and L. Harrington. 1998. Reconstitution of human telomerase activity in vitro. Curr. Biol. 8:177-180. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources