

BMPER, a novel endothelial cell precursor-derived protein, antagonizes bone morphogenetic protein signaling and endothelial cell differentiation
- PMID: 12897139
- PMCID: PMC166349
- DOI: 10.1128/MCB.23.16.5664-5679.2003
BMPER, a novel endothelial cell precursor-derived protein, antagonizes bone morphogenetic protein signaling and endothelial cell differentiation
Abstract
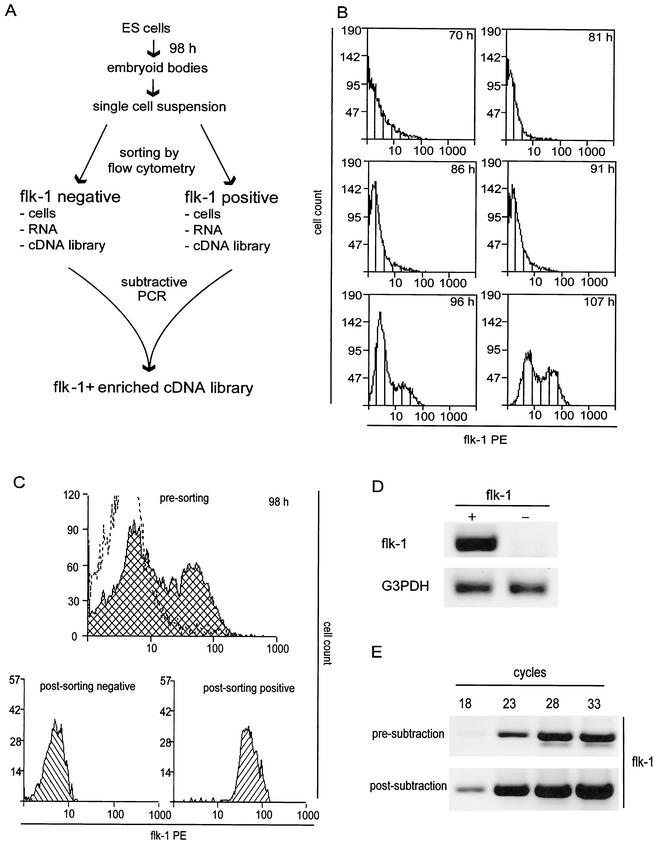
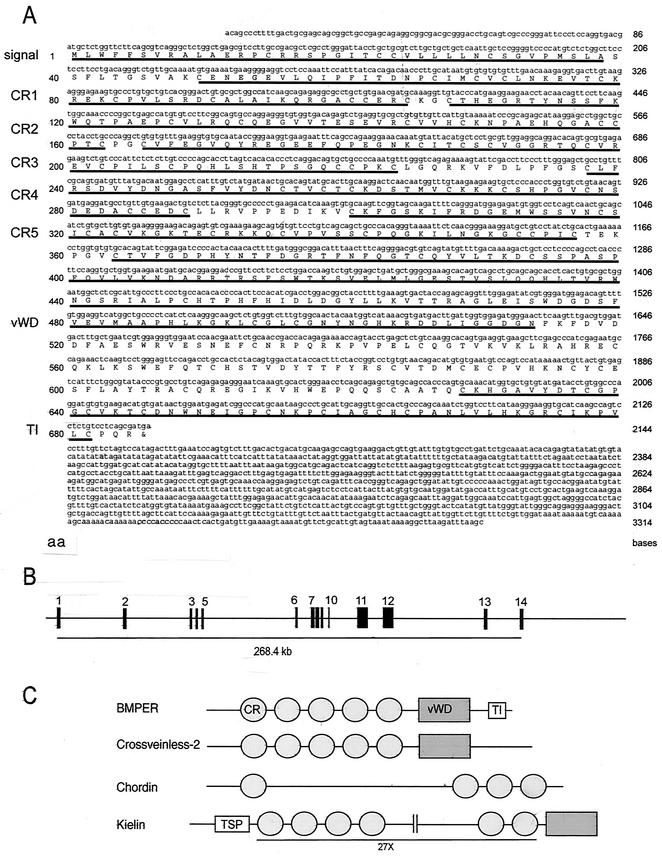
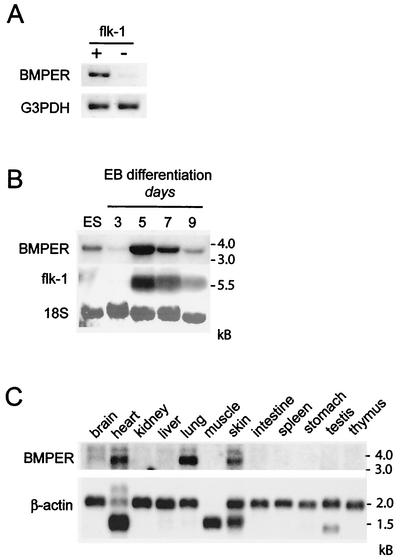
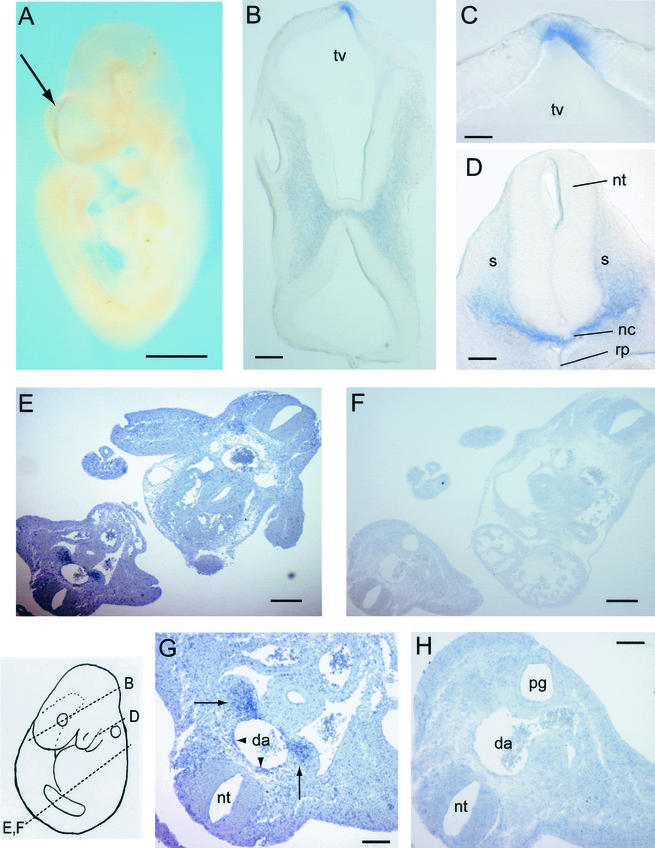
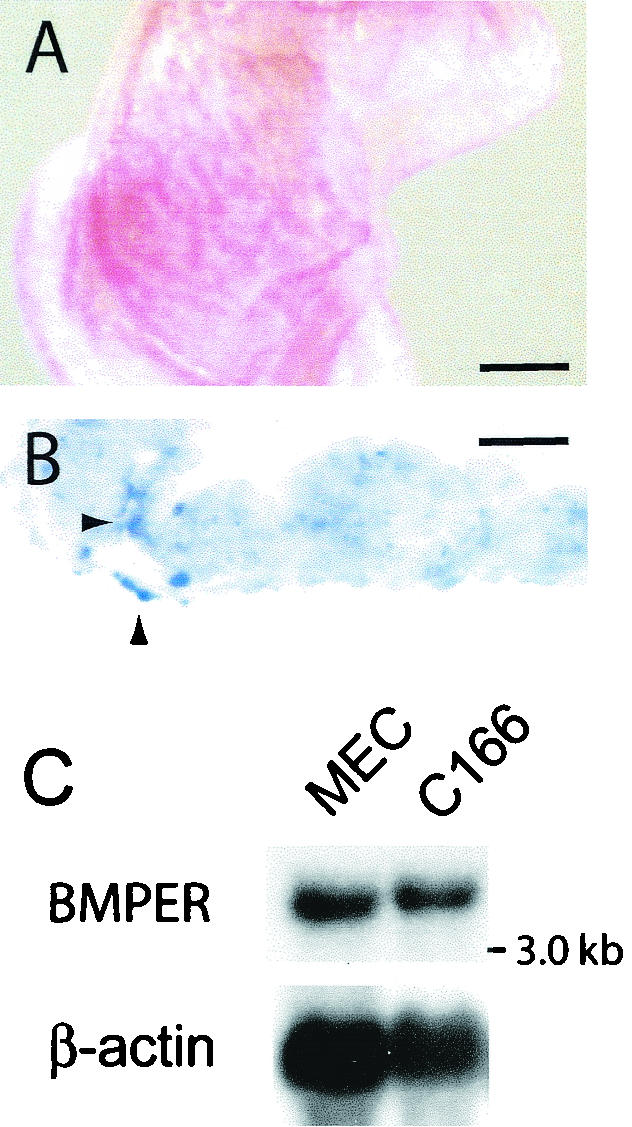
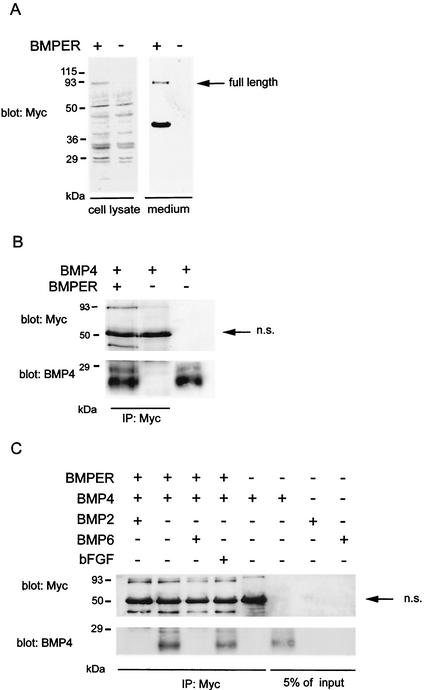
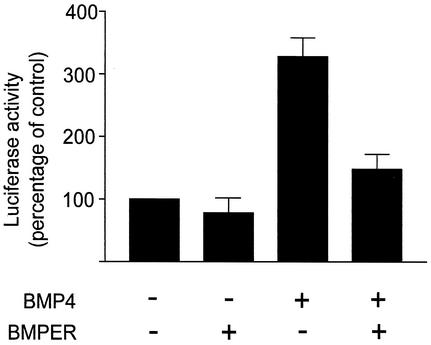
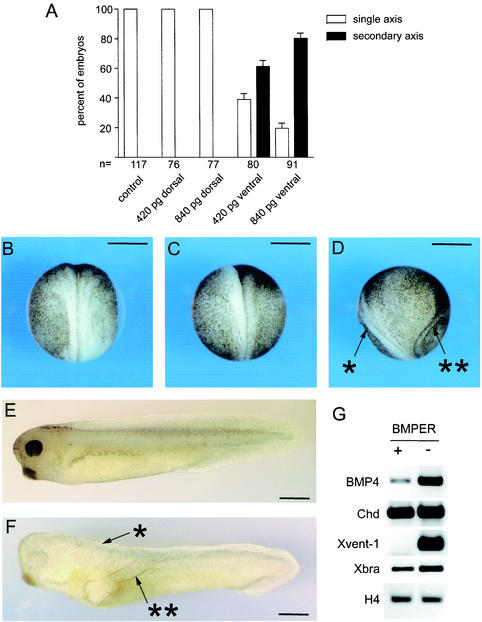
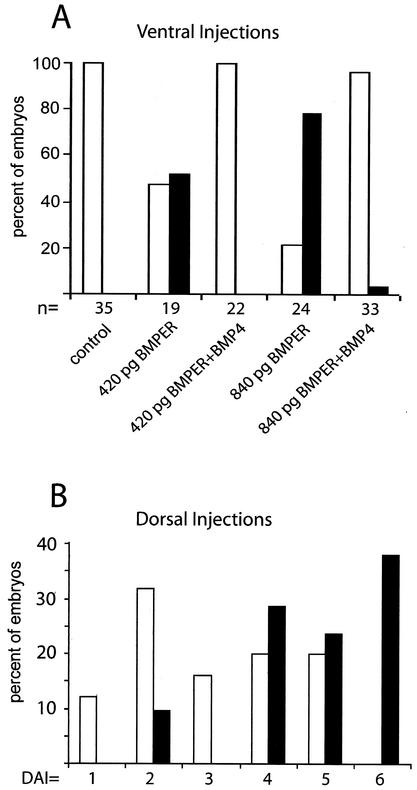
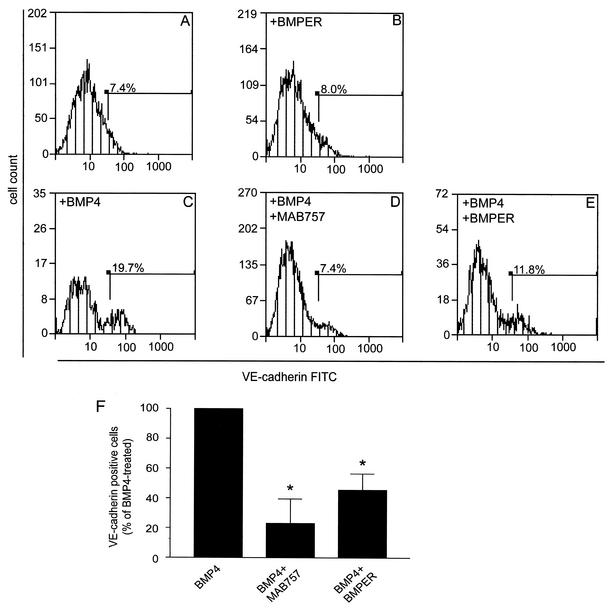
The development of endothelial cell precursors is essential for vasculogenesis. We screened for differentially expressed transcripts in endothelial cell precursors in developing mouse embryoid bodies. We cloned a complete cDNA encoding a protein that contains an amino-terminal signal peptide, five cysteine-rich domains, a von Willebrand D domain, and a trypsin inhibitor domain. We termed this protein BMPER (bone morphogenetic protein [BMP]-binding endothelial cell precursor-derived regulator). BMPER is specifically expressed in flk-1-positive cells and parallels the time course of flk-1 induction in these cells. In situ hybridization in mouse embryos demonstrates dorsal midline staining and staining of the aorto-gonadal-mesonephric region, which is known to host vascular precursor cells. BMPER is a secreted protein that directly interacts with BMP2, BMP4, and BMP6 and antagonizes BMP4-dependent Smad5 activation. In Xenopus embryos, ventral injection of BMPER mRNA results in axis duplication and downregulation of the expression of Xvent-1 (downstream target of Smad signaling). In an embryoid body differentiation assay, BMP4-dependent differentiation of endothelial cells in embryoid bodies is also antagonized by BMPER. Taken together, our data indicate that BMPER is a novel BMP-binding protein that is expressed by endothelial cell precursors, has BMP-antagonizing activity, and may play a role in endothelial cell differentiation by modulating local BMP activity.
Figures
References
-
- Adelman, C. A., S. Chattopadhyay, and J. J. Bieker. 2002. The BMP/BMPR/Smad pathway directs expression of the erythroid-specific EKLF and GATA1 transcription factors during embryoid body differentiation in serum-free media. Development 129:539-549. - PubMed
-
- Allen, S., A. M. Abuzenadah, J. L. Blagg, J. Hinks, I. M. Nesbitt, A. C. Goodeve, T. Gursel, J. Ingerslev, I. R. Peake, and M. E. Daly. 2000. Two novel type 2N von Willebrand disease-causing mutations that result in defective factor VIII binding, multimerization, and secretion of von Willebrand factor. Blood 95:2000-2007. - PubMed
-
- Balemans, W., and W. vanHul. 2002. Extracellular regulation of BMP signaling in vertebrates: a cocktail of modulators. Dev. Biol. 250:231-250. - PubMed
-
- Bautch, V. 2002. Embryonic stem cell differentiation and the vascular lineage. Humana Press, Totowa, N.J. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases