

HoxB5 is an upstream transcriptional switch for differentiation of the vascular endothelium from precursor cells
- PMID: 12897140
- PMCID: PMC166331
- DOI: 10.1128/MCB.23.16.5680-5691.2003
HoxB5 is an upstream transcriptional switch for differentiation of the vascular endothelium from precursor cells
Abstract
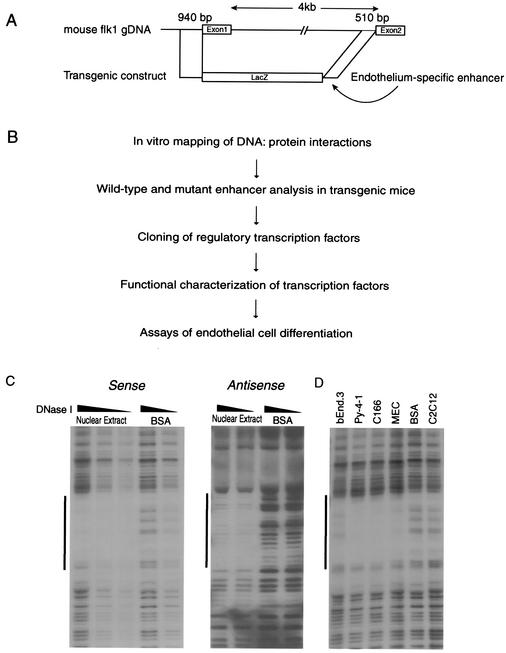
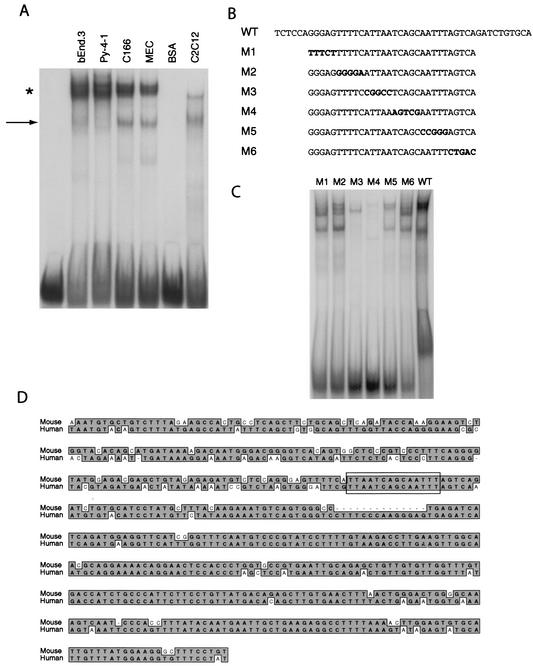
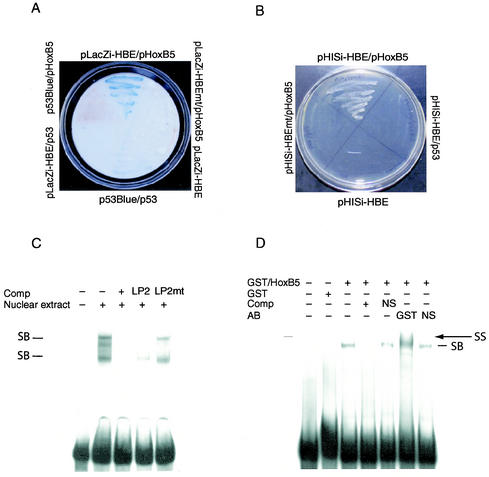
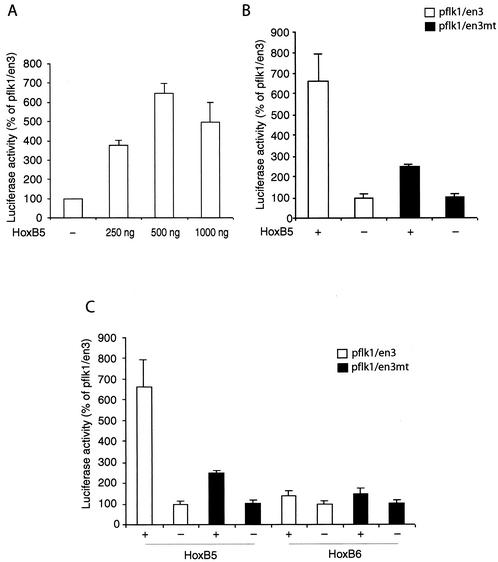
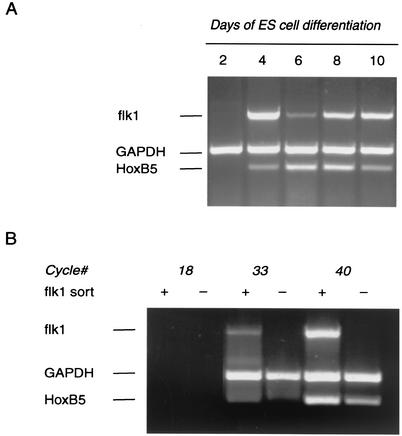
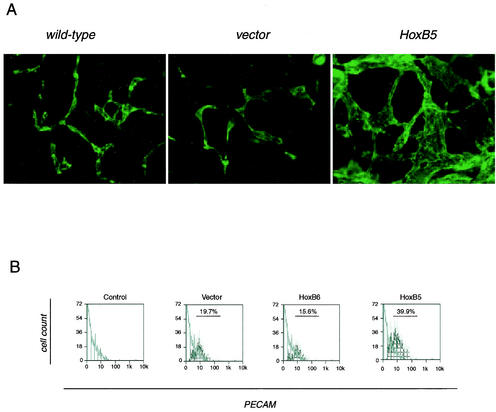
Endothelial cells differentiate from mesoderm-derived precursors to initiate the earliest events in vascular development. Although the signaling events that regulate the successive steps of vascular development are known in some detail, the transcriptional processes that regulate the first steps in vasculogenesis are not well defined. We have studied the regulatory mechanisms of flk1 expression as a model to understand the upstream events in endothelial cell differentiation, since flk1 is the earliest marker of endothelial precursors. Using a variety of biochemical approaches, we identified a cis-acting element in the first intron of the flk1 gene that is required for endothelium-dependent expression in transgenic reporter gene assays. Using the yeast one-hybrid system, we identified HoxB5 as the transcription factor that binds this cis-acting element, the HoxB5-binding element (HBE). HoxB5 mRNA colocalized with flk1 expression in differentiating embryoid bodies, and HoxB5 potently transactivated the flk1 promoter in an HBE-dependent fashion in transient-transfection assays. Overexpression of HoxB5 led to expansion of flk1(+) angioblasts in differentiating embryoid bodies and increased the number of PECAM (platelet-endothelial cell adhesion molecule)-positive primitive blood vessels. HoxB5 is necessary and sufficient to activate the cell-intrinsic events that regulate the differentiation of angioblasts and mature endothelial cells from their mesoderm-derived precursors.
Figures


Similar articles
-
Pdgfrα and Flk1 are direct target genes of Mixl1 in differentiating embryonic stem cells.Stem Cell Res. 2012 Mar;8(2):165-79. doi: 10.1016/j.scr.2011.09.007. Epub 2011 Oct 8. Stem Cell Res. 2012. PMID: 22265737
-
NANOG induction of fetal liver kinase-1 (FLK1) transcription regulates endothelial cell proliferation and angiogenesis.Blood. 2011 Feb 3;117(5):1761-9. doi: 10.1182/blood-2010-07-295261. Epub 2010 Nov 30. Blood. 2011. PMID: 21119109 Free PMC article.
-
Etv2/ER71 induces vascular mesoderm from Flk1+PDGFRα+ primitive mesoderm.Blood. 2011 Dec 22;118(26):6975-86. doi: 10.1182/blood-2011-05-352658. Epub 2011 Sep 12. Blood. 2011. PMID: 21911838
-
Vasculogenesis.Annu Rev Cell Dev Biol. 1995;11:73-91. doi: 10.1146/annurev.cb.11.110195.000445. Annu Rev Cell Dev Biol. 1995. PMID: 8689573 Review.
-
Transcriptional control of endothelial cell development.Dev Cell. 2009 Feb;16(2):180-95. doi: 10.1016/j.devcel.2009.01.014. Dev Cell. 2009. PMID: 19217421 Free PMC article. Review.
Cited by
-
The dual roles of homeobox genes in vascularization and wound healing.Cell Adh Migr. 2012 Nov-Dec;6(6):457-70. doi: 10.4161/cam.22164. Epub 2012 Oct 17. Cell Adh Migr. 2012. PMID: 23076135 Free PMC article. Review.
-
HOXA9 methylation by PRMT5 is essential for endothelial cell expression of leukocyte adhesion molecules.Mol Cell Biol. 2012 Apr;32(7):1202-13. doi: 10.1128/MCB.05977-11. Epub 2012 Jan 23. Mol Cell Biol. 2012. PMID: 22269951 Free PMC article.
-
HOX Genes Family and Cancer: A Novel Role for Homeobox B9 in the Resistance to Anti-Angiogenic Therapies.Cancers (Basel). 2020 Nov 8;12(11):3299. doi: 10.3390/cancers12113299. Cancers (Basel). 2020. PMID: 33171691 Free PMC article. Review.
-
Analysis of homeobox gene action may reveal novel angiogenic pathways in normal placental vasculature and in clinical pregnancy disorders associated with abnormal placental angiogenesis.Front Pharmacol. 2014 Jun 4;5:133. doi: 10.3389/fphar.2014.00133. eCollection 2014. Front Pharmacol. 2014. PMID: 24926269 Free PMC article. Review.
-
Unraveling the transcriptional determinants of liver sinusoidal endothelial cell specialization.Am J Physiol Gastrointest Liver Physiol. 2020 Apr 1;318(4):G803-G815. doi: 10.1152/ajpgi.00215.2019. Epub 2020 Mar 2. Am J Physiol Gastrointest Liver Physiol. 2020. PMID: 32116021 Free PMC article.
References
-
- Aitsebaomo, J., M. Kingsley-Kallesen, Y. Wu, T. Quertermous, and C. Patterson. 2001. Vezf1/DB1 is an endothelial cell-specific transcription factor that regulates expression of the endothelin-1 promoter. J. Biol. Chem. 276:39197-39205. - PubMed
-
- Ambler, C. A., J. L. Nowicki, A. C. Burke, and V. L. Bautch. 2001. Assembly of trunk and limb blood vessels involves extensive migration and vasculogenesis of somite-derived angioblasts. Dev. Biol. 234:352-364. - PubMed
-
- Antonchuk, J., G. Sauvageau, and R. K. Humphries. 2002. HOXB4-induced expansion of adult hematopoietic stem cells ex vivo. Cell 109:39-45. - PubMed
-
- Bautch, V., W. Stanford, R. Rapoport, S. Russell, R. Byrum, and T. Futch. 1996. Blood island formation in atached culture of murine embryonic stem cells. Dev. Dyn. 205:1-12. - PubMed
-
- Bautch, V. L. 2002. Embryonic stem cell differentiation and the vascular lineage. Methods Mol. Biol. 185:117-125. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases