

Promoter-specific roles for liver X receptor/corepressor complexes in the regulation of ABCA1 and SREBP1 gene expression
- PMID: 12897148
- PMCID: PMC166346
- DOI: 10.1128/MCB.23.16.5780-5789.2003
Promoter-specific roles for liver X receptor/corepressor complexes in the regulation of ABCA1 and SREBP1 gene expression
Abstract
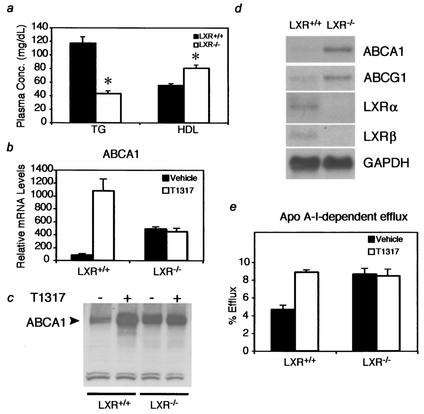
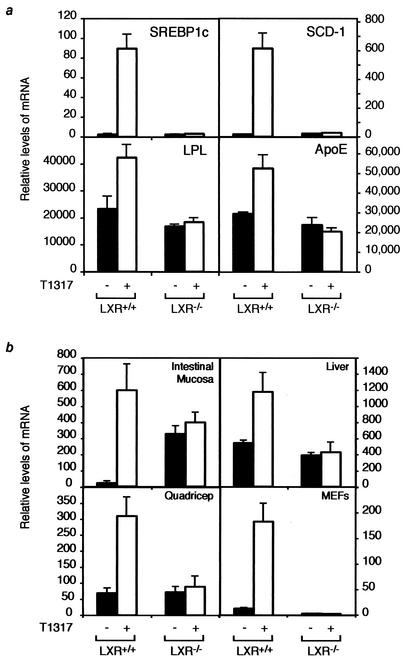
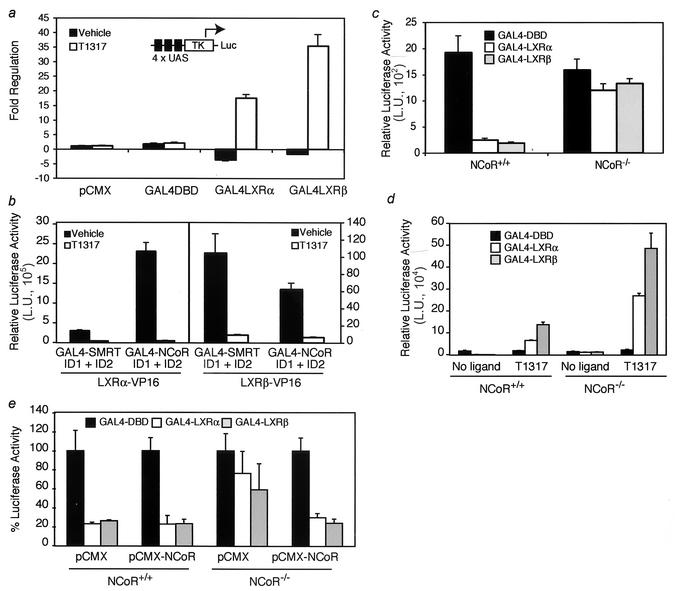
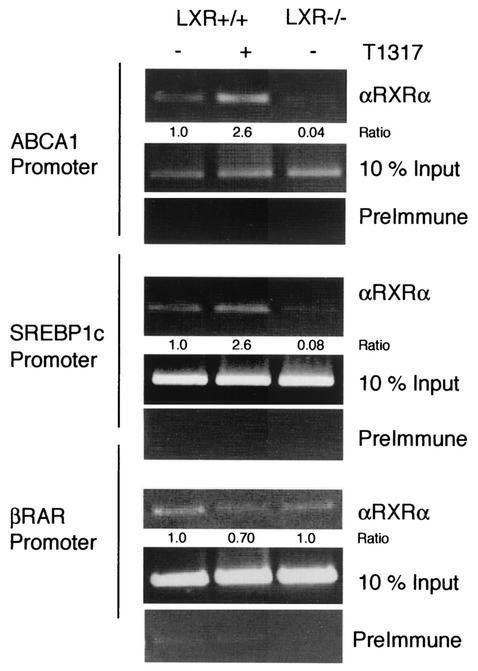
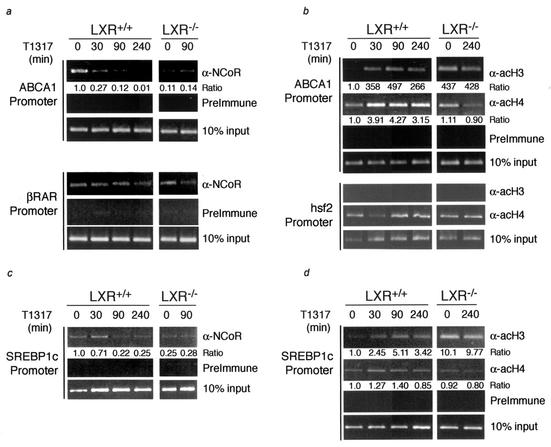
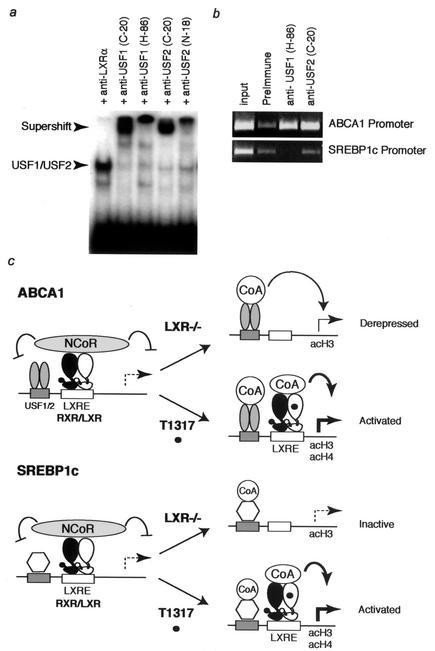
Liver X receptors (LXRs) regulate the expression of genes involved in cholesterol and fatty acid homeostasis, including the genes for ATP-binding cassette transporter A1 (ABCA1) and sterol response element binding protein 1 (SREBP1). Loss of LXR leads to derepression of the ABCA1 gene in macrophages and the intestine, while the SREBP1c gene remains transcriptionally silent. Here we report that high-density-lipoprotein (HDL) cholesterol levels are increased in LXR-deficient mice, suggesting that derepression of ABCA1 and possibly other LXR target genes in selected tissues is sufficient to result in enhanced HDL biogenesis at the whole-body level. We provide several independent lines of evidence indicating that the repressive actions of LXRs are dependent on interactions with the nuclear receptor corepressor (NCoR) and the silencing mediator of retinoic acid and thyroid hormone receptors (SMRT). While dissociation of NCoR and SMRT results in derepression of the ABCA1 gene in macrophages, it is not sufficient for derepression of the SREBP1c gene. These findings reveal differential requirements for corepressors in the regulation of genes involved in cholesterol and fatty acid homeostasis and raise the possibility that these interactions may be exploited to develop synthetic ligands that selectively modulate LXR actions in vivo.
Figures
References
-
- Chen, J. D., and R. M. Evans. 1995. A transcriptional co-repressor that interacts with nuclear hormone receptors. Nature 377:454-457. - PubMed
-
- Claudel, T., M. D. Leibowitz, C. Fievet, A. Tailleux, B. Wagner, J. J. Repa, G. Torpier, J. M. Lobaccaro, J. R. Paterniti, D. J. Mangelsdorf, R. A. Heyman, and J. Auwerx. 2001. Reduction of atherosclerosis in apolipoprotein E knockout mice by activation of the retinoid X receptor. Proc. Natl. Acad. Sci. USA 98:2610-2615. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials