

An Arabidopsis mutant resistant to thaxtomin A, a cellulose synthesis inhibitor from Streptomyces species
- PMID: 12897252
- PMCID: PMC167169
- DOI: 10.1105/tpc.013342
An Arabidopsis mutant resistant to thaxtomin A, a cellulose synthesis inhibitor from Streptomyces species
Abstract
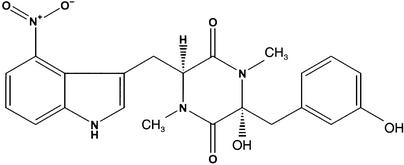
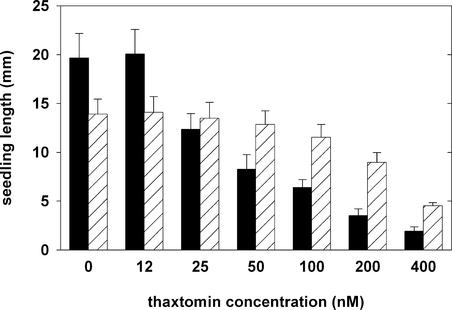
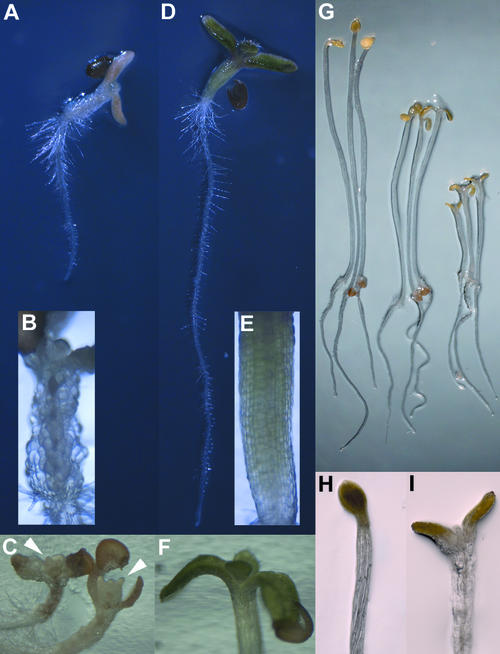
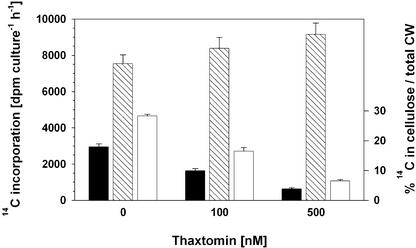
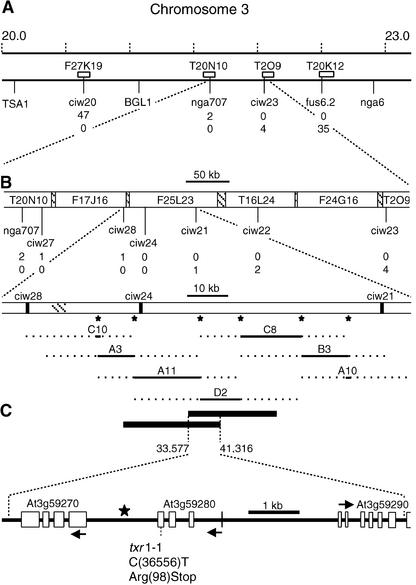
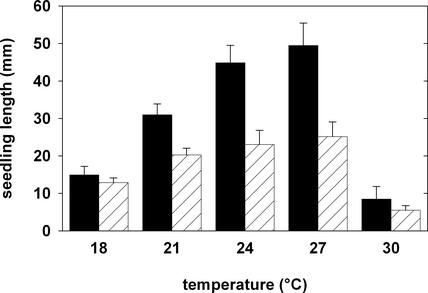
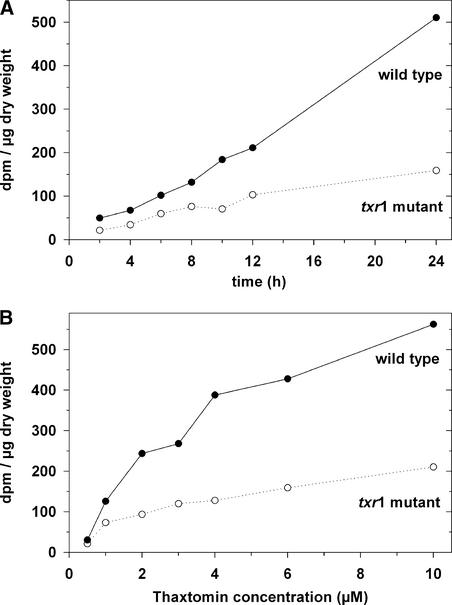
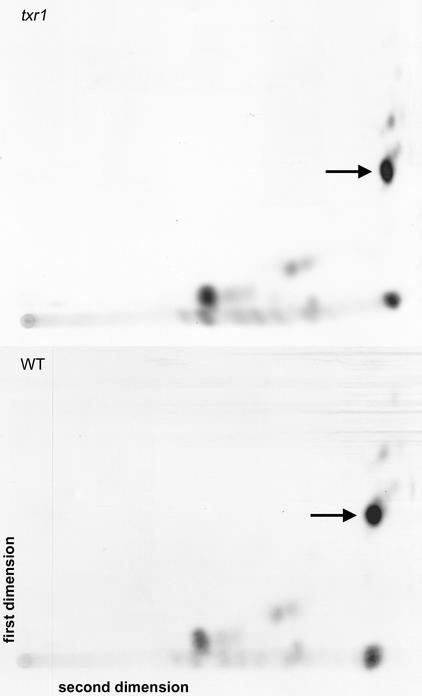
Thaxtomin A is a phytotoxin produced by Streptomyces scabies and other Streptomyces species, the causative agents of common scab disease in potato and other taproot crops. At nanomolar concentrations, thaxtomin causes dramatic cell swelling, reduced seedling growth, and inhibition of cellulose synthesis in Arabidopsis. We identified a mutant of Arabidopsis, designated txr1, that exhibits increased resistance to thaxtomin as a result of a decrease in the rate of toxin uptake. The TXR1 gene was identified by map-based cloning and found to encode a novel, small protein with no apparent motifs or organelle-targeting signals. The protein, which has homologs in all fully sequenced eukaryotic genomes, is expressed in all tissues and during all developmental stages analyzed. Microarray transcript profiling of some 14,300 genes revealed two stomatin-like genes that were expressed differentially in the txr1 mutant and the wild type. We propose that TXR1 is a regulator of a transport mechanism.
Figures



References
-
- Acuna, I.A., Strobel, G.A., Jacobsen, B.L., and Corsini, D.L. (2001). Glucosylation as a mechanism of resistance to thaxtomin A in potatoes. Plant Sci. 161, 77–88.
-
- Anzai, H., Yoneyama, K., and Yamagushi, I. (1989). Transgenic tobacco resistant to a bacterial disease by the detoxification of a pathogenic toxin. Mol. Gen. Genet. 219, 492–494.
-
- Clough, S.J., and Bent, A.F. (1998). Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 16, 735–743. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
