

Arabidopsis AtGPAT1, a member of the membrane-bound glycerol-3-phosphate acyltransferase gene family, is essential for tapetum differentiation and male fertility
- PMID: 12897259
- PMCID: PMC167176
- DOI: 10.1105/tpc.012427
Arabidopsis AtGPAT1, a member of the membrane-bound glycerol-3-phosphate acyltransferase gene family, is essential for tapetum differentiation and male fertility
Abstract
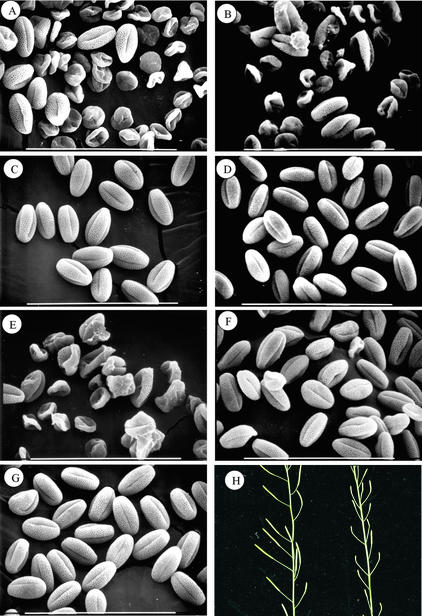
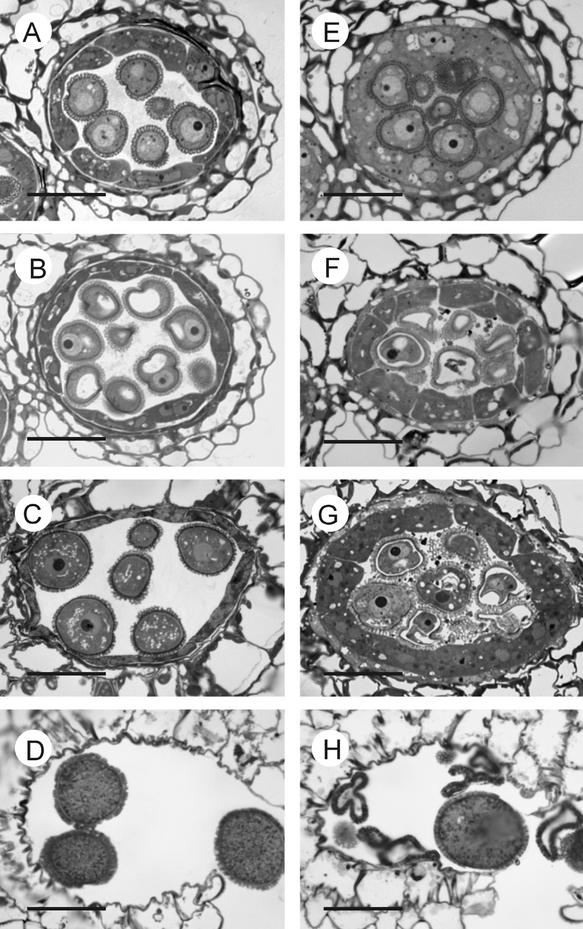
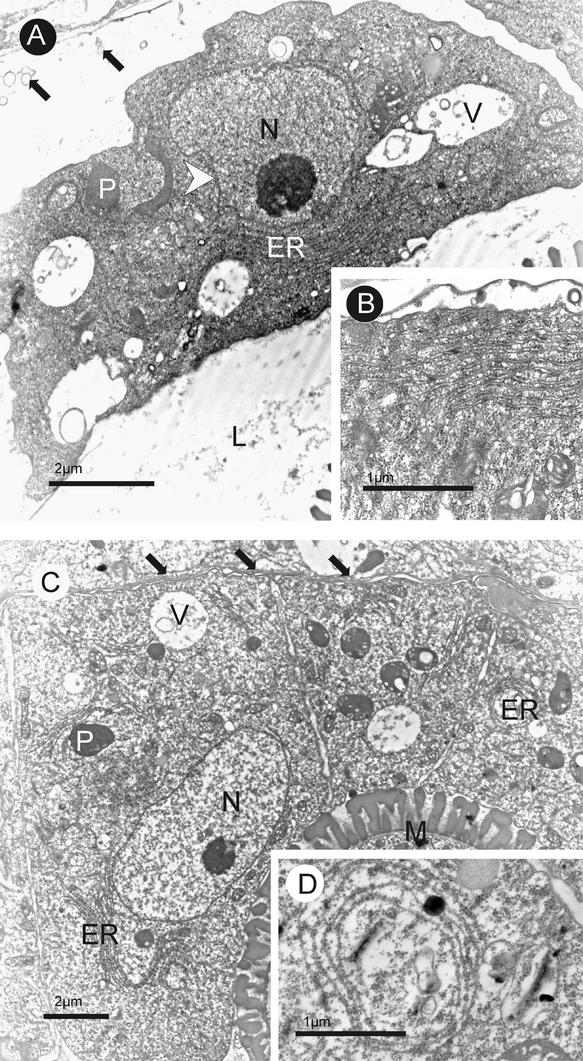
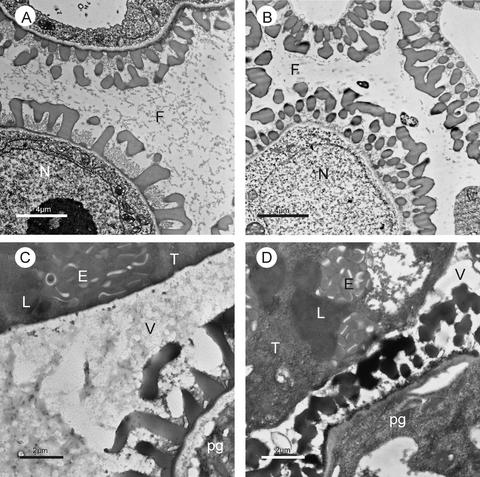
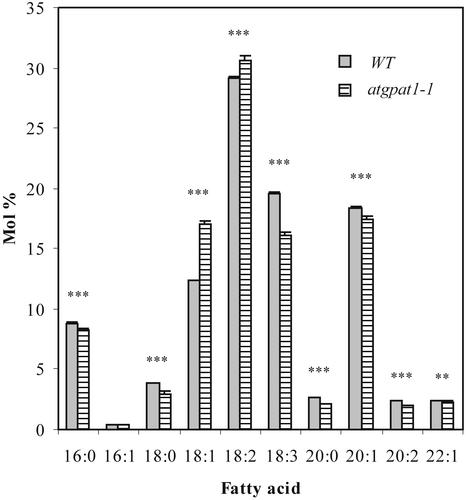
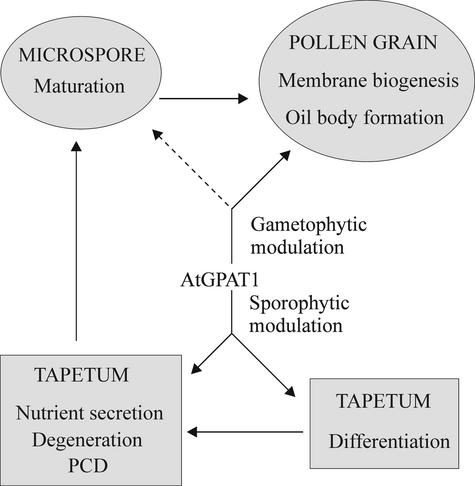
Membrane-bound glycerol-3-phosphate acyltransferase (GPAT; EC 2.3.1.15) mediates the initial step of glycerolipid biosynthesis in the extraplastidic compartments of plant cells. Here, we report the molecular characterization of a novel GPAT gene family from Arabidopsis, designated AtGPAT. The corresponding polypeptides possess transmembrane domains and GPAT activity when expressed heterologously in a yeast lipid mutant. The functional significance of one isoform, AtGPAT1, is the focus of the present study. Disruption of the AtGPAT1 gene causes a massive pollen development arrest, and subsequent introduction of the gene into the mutant plant rescues the phenotype, illustrating a pivotal role for AtGPAT1 in pollen development. Microscopic examinations revealed that the gene lesion results in a perturbed degeneration of the tapetum, which is associated with altered endoplasmic reticulum profiles and reduced secretion. In addition to the sporophytic effect, AtGPAT1 also exerts a gametophytic effect on pollen performance, as the competitive ability of a pollen grain to pollinate is dependent on the presence of an AtGPAT1 gene. Deficiency in AtGPAT1 correlates with several fatty acid composition changes in flower tissues and seeds. Unexpectedly, however, a loss of AtGPAT1 causes no significant change in seed oil content.
Figures

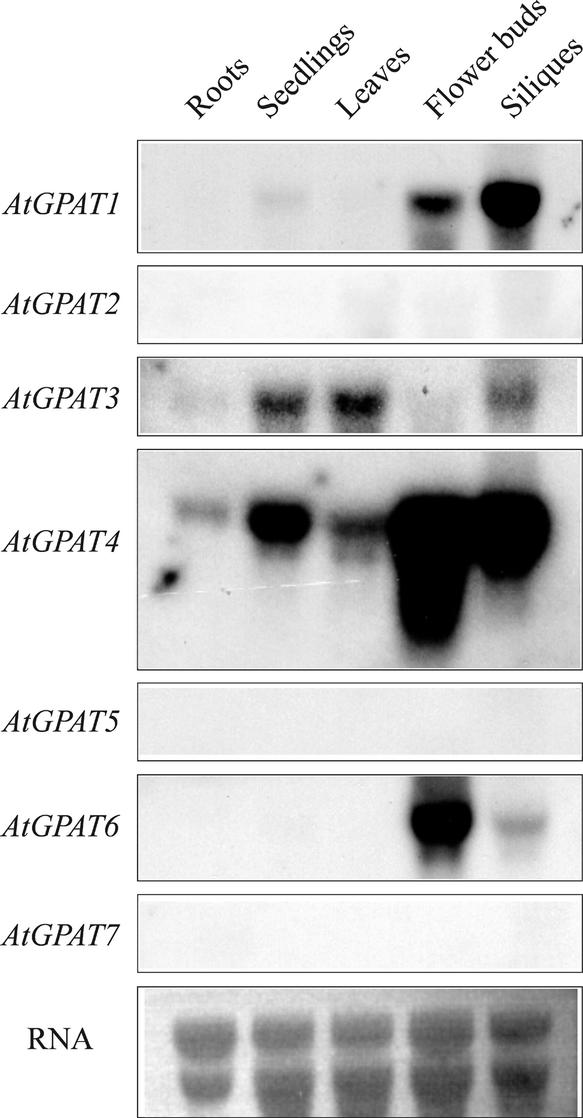
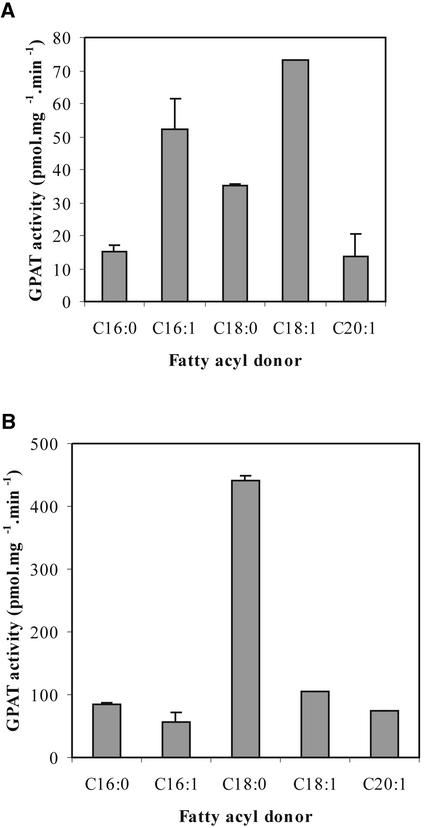
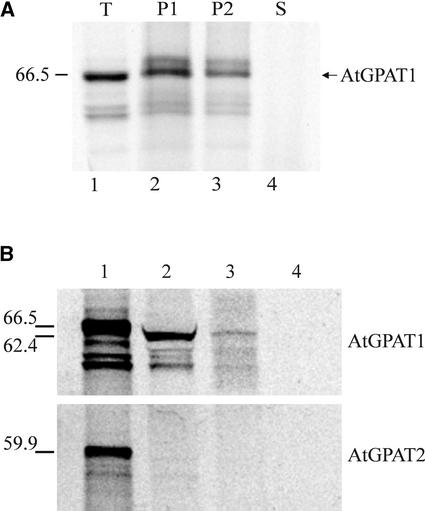
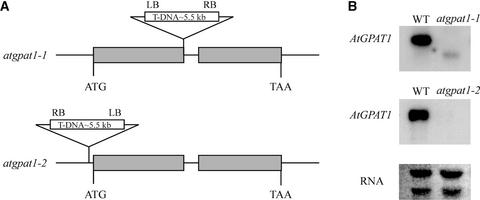

References
-
- Altschul, S.F., Gish, W., Miller, W., Myers, E.W., and Lipman, D.J. (1990). Basic local alignment search tool. J. Mol. Biol. 215, 403–410. - PubMed
-
- Athenstaedt, K., and Daum, G. (1999). Phosphatidic acid, a key intermediate in lipid metabolism. Eur. J. Biochem. 266, 1–16. - PubMed
-
- Bafor, M., Stobart, A.K., and Stymne, S. (1990). Properties of the glycerol acylating enzyme in microsomal preparations from the developing seeds of safflower (Carthamus tinctorius) and turnip rape (Brassica campestris) and their ability to assemble cocoa-butter type fats. J. Am. Oil Chem. Soc. 67, 217–225.
-
- Bligh, E.G., and Dyer, W.J. (1959). A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 37, 911–917. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
