

Auditory cortical responses elicited in awake primates by random spectrum stimuli
- PMID: 12904480
- PMCID: PMC1945239
- DOI: 10.1523/JNEUROSCI.23-18-07194.2003
Auditory cortical responses elicited in awake primates by random spectrum stimuli
Abstract
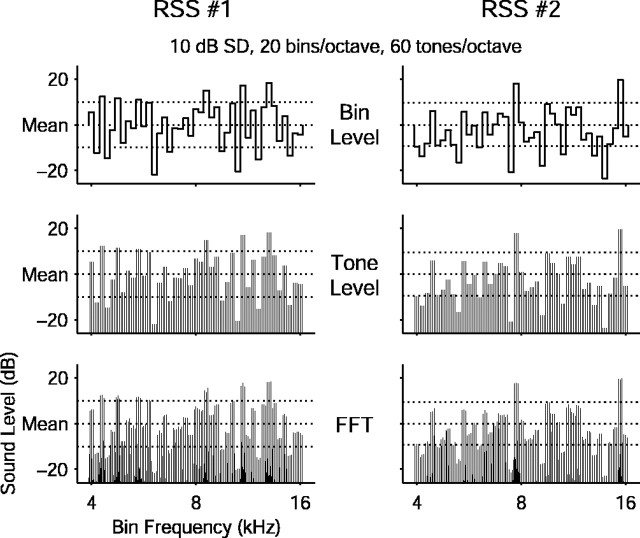
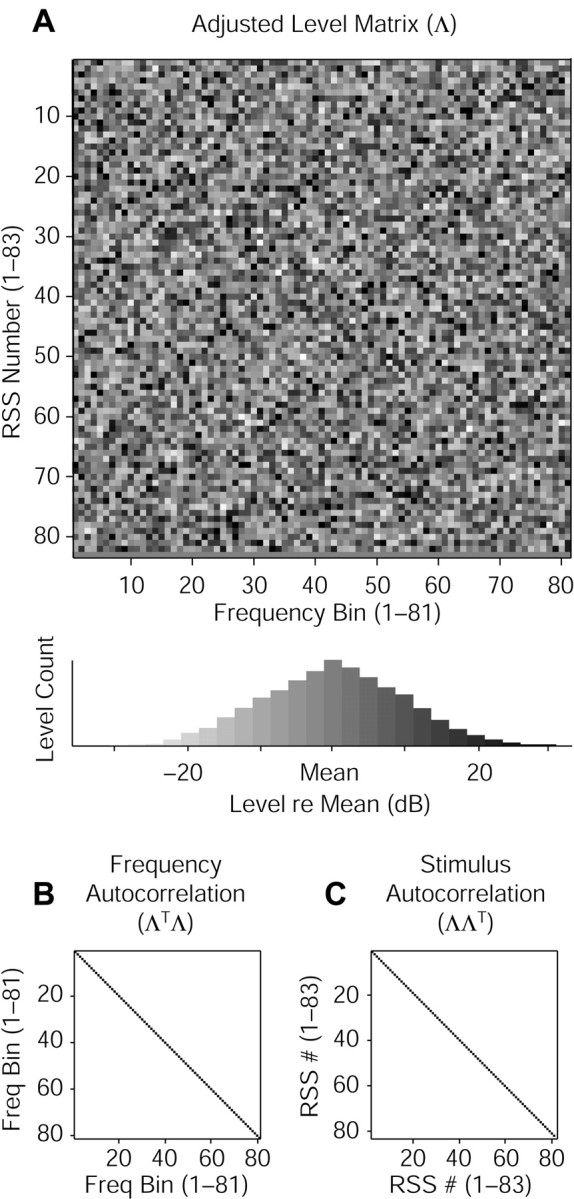
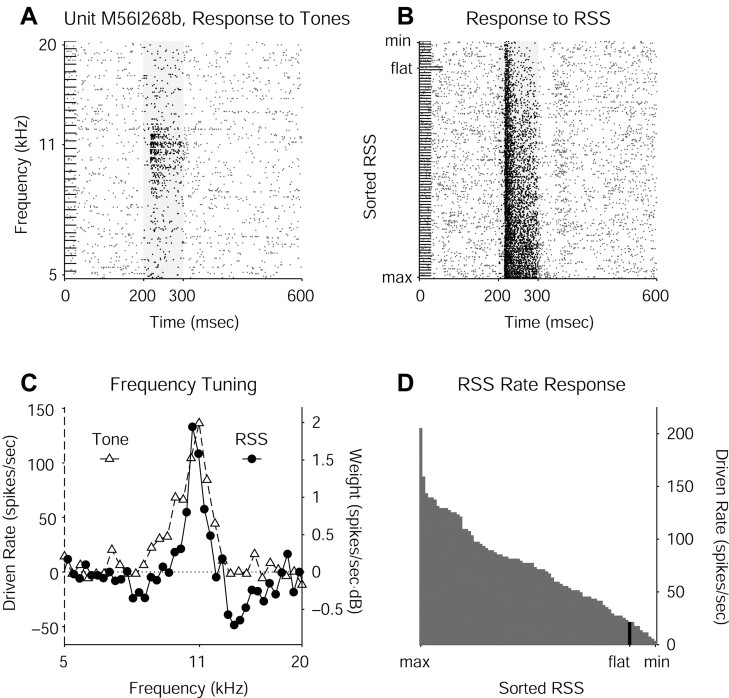
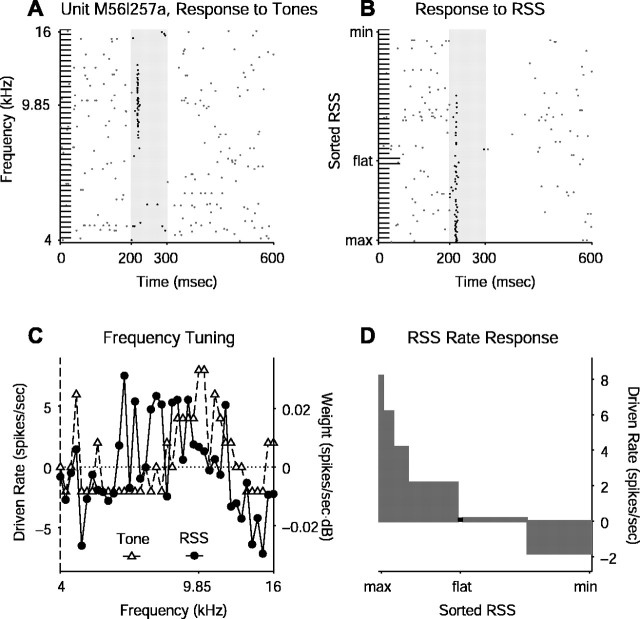
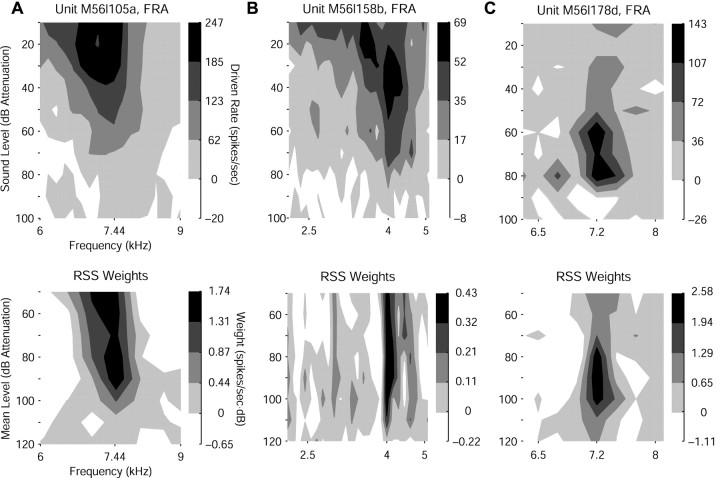
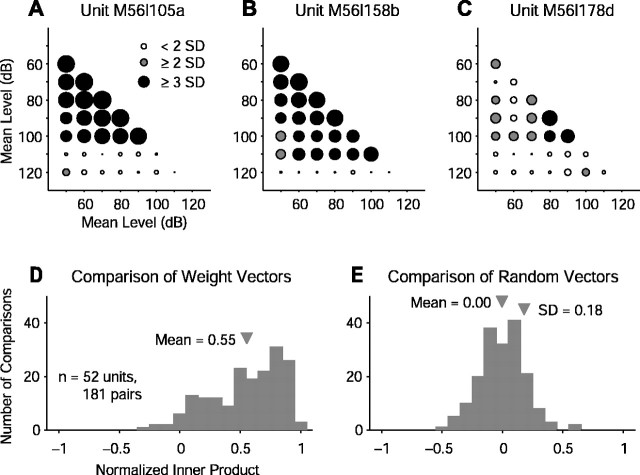
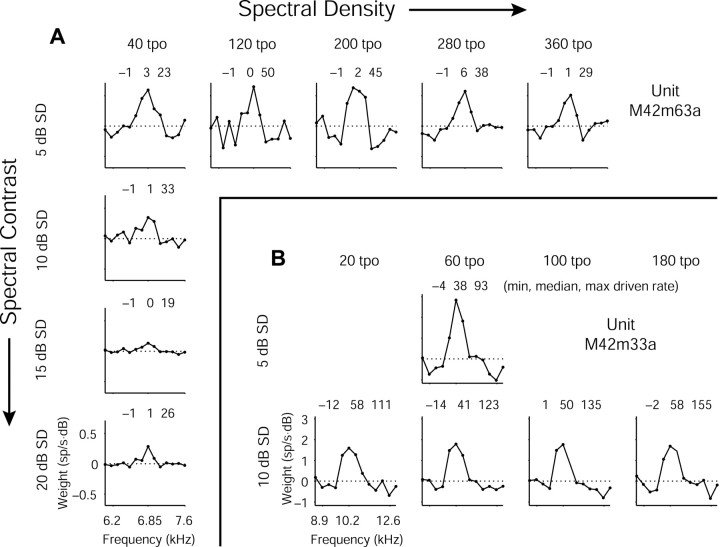
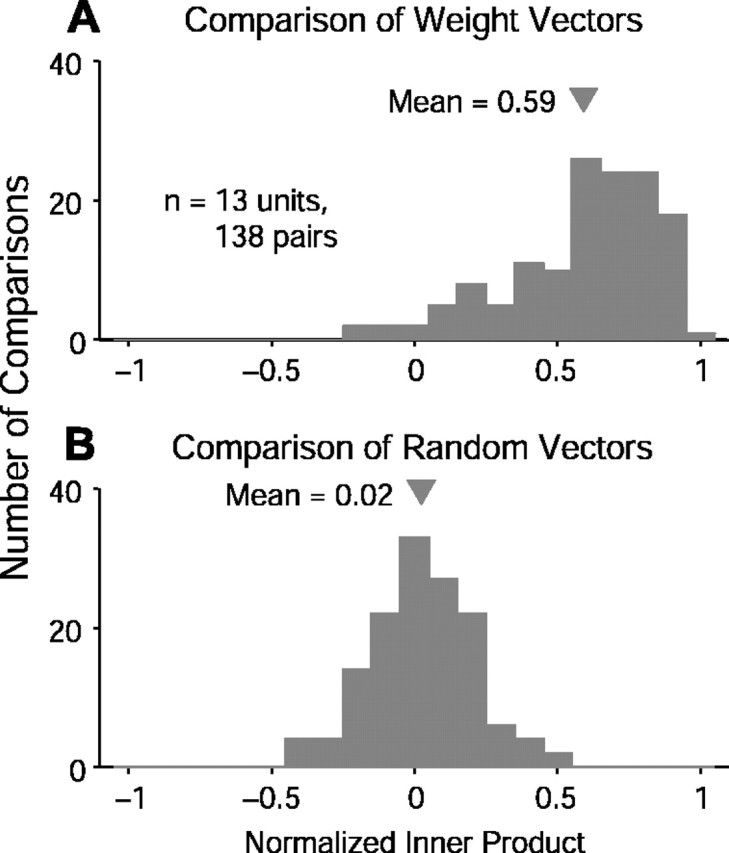
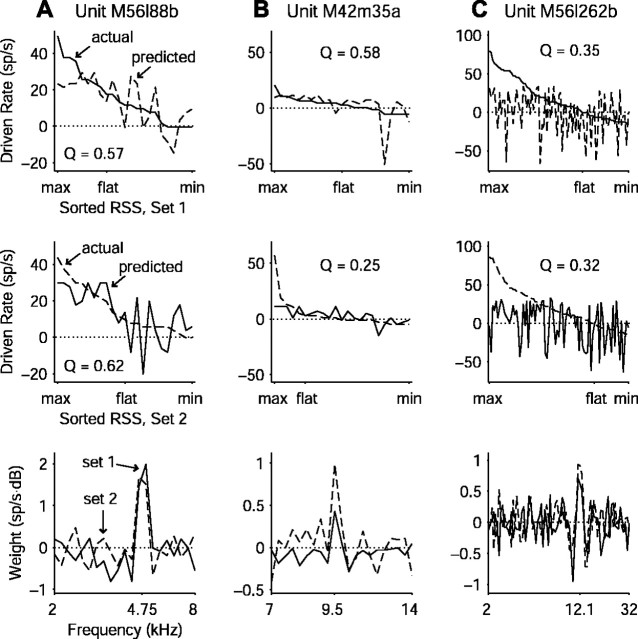
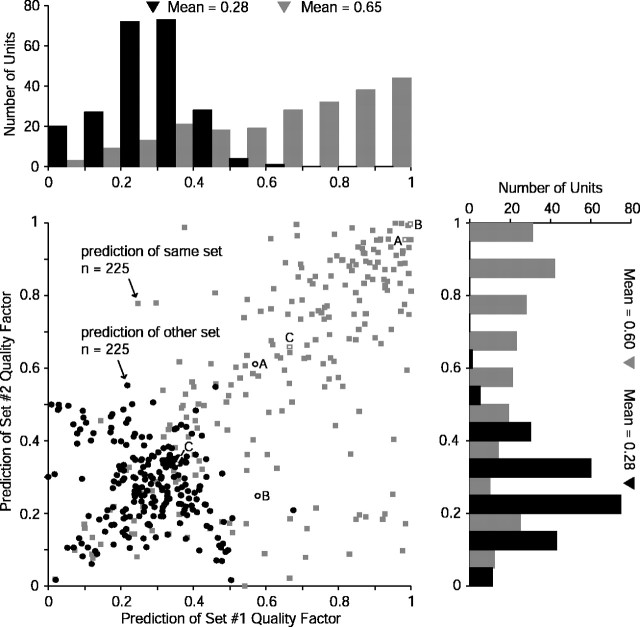
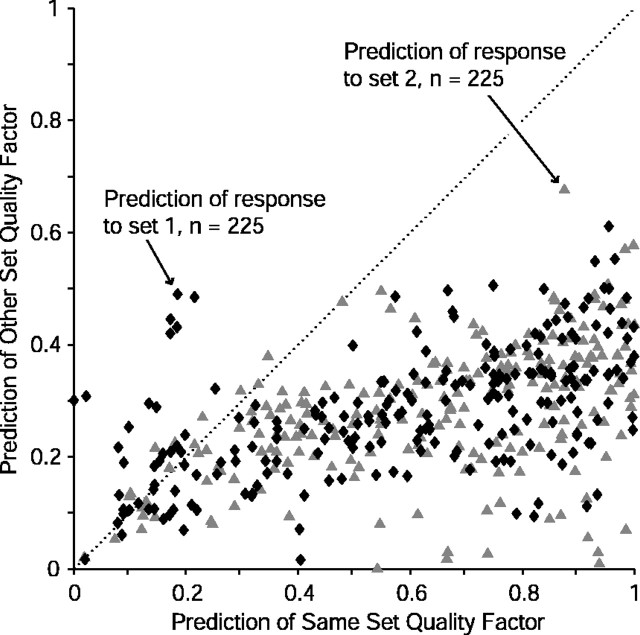
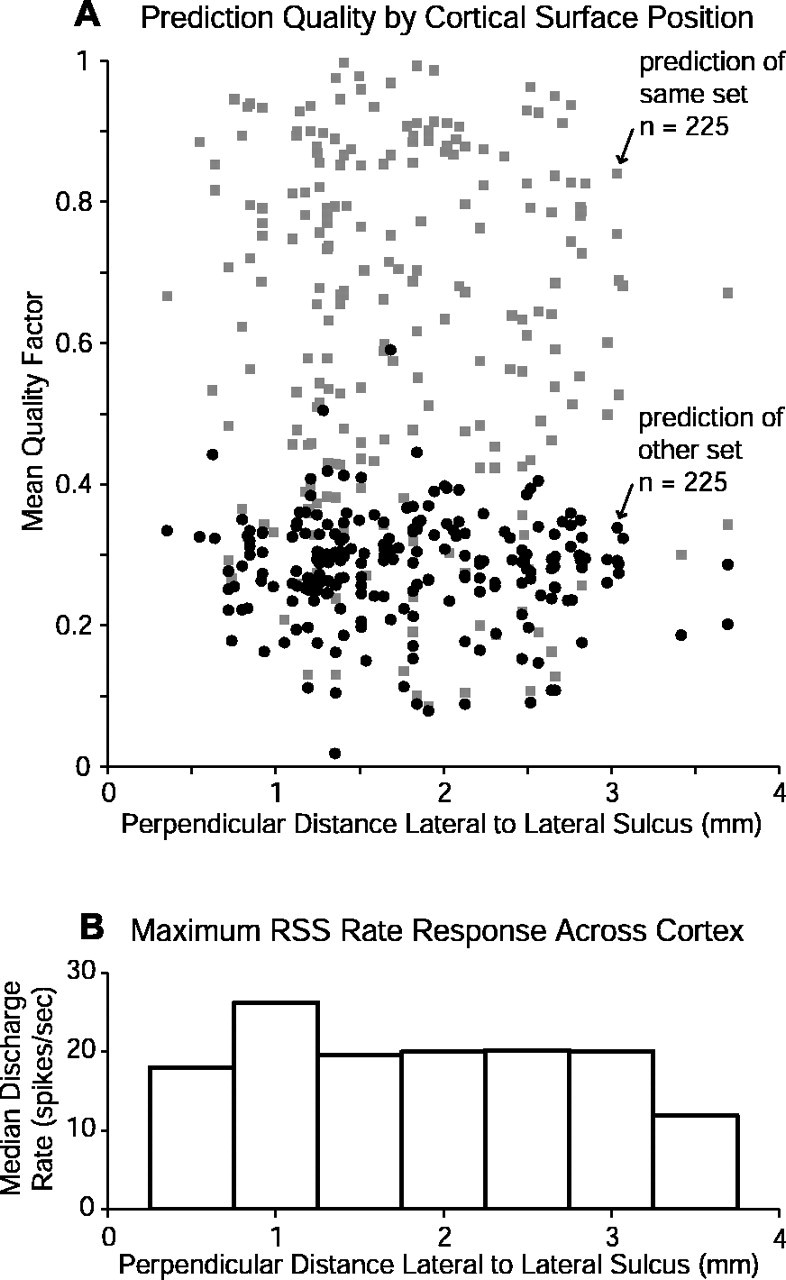
Contrary to findings in subcortical auditory nuclei, auditory cortex neurons have traditionally been described as spiking only at the onsets of simple sounds such as pure tones or bandpass noise and to acoustic transients in complex sounds. Furthermore, primary auditory cortex (A1) has traditionally been described as mostly tone responsive and the lateral belt area of primates as mostly noise responsive. The present study was designed to unify the study of these two cortical areas using random spectrum stimuli (RSS), a new class of parametric, wideband, stationary acoustic stimuli. We found that 60% of all neurons encountered in A1 and the lateral belt of awake marmoset monkeys (Callithrix jacchus) showed significant changes in firing rates in response to RSS. Of these, 89% showed sustained spiking in response to one or more individual RSS, a substantially greater percentage than would be expected from traditional studies, indicating that RSS are well suited for studying these two cortical areas. When firing rates elicited by RSS were used to construct linear estimates of frequency tuning for these sustained responders, the shape of the estimate function remained relatively constant throughout the stimulus interval and across the stimulus properties of mean sound level, spectral density, and spectral contrast. This finding indicates that frequency tuning computed from RSS reflects a robust estimate of the actual tuning of a neuron. Use of this estimate to predict rate responses to other RSS, however, yielded poor results, implying that auditory cortex neurons integrate information across frequency nonlinearly. No systematic difference in prediction quality between A1 and the lateral belt could be detected.
Figures

References
-
- Abeles M, Goldstein Jr MH ( 1970) Functional architecture in cat primary auditory cortex: columnar organization and organization according to depth. J Neurophysiol 33: 172-187. - PubMed
-
- Abeles M, Goldstein Jr MH ( 1972) Responses of single units in the primary auditory cortex of the cat to tones and to tone pairs. Brain Res 42: 337-352. - PubMed
-
- Aertsen AM, Johannesma PI ( 1981) The spectro-temporal receptive field. A functional characteristic of auditory neurons. Biol Cybern 42: 133-143. - PubMed
-
- Anderson JS, Lampl I, Gillespie DC, Ferster D ( 2000) The contribution of noise to contrast invariance of orientation tuning in cat visual cortex. Science 290: 1968-1972. - PubMed
-
- Barbour DL, Wang X ( 2002) Temporal coherence sensitivity in auditory cortex. J Neurophysiol 88: 2684-2699. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources