

Neuronal populations and single cells representing learned auditory objects
- PMID: 12904792
- PMCID: PMC2631575
- DOI: 10.1038/nature01731
Neuronal populations and single cells representing learned auditory objects
Abstract
The neural representations associated with learned auditory behaviours, such as recognizing individuals based on their vocalizations, are not well described. Higher vertebrates learn to recognize complex conspecific vocalizations that comprise sequences of easily identified, naturally occurring auditory objects, which should facilitate the analysis of higher auditory pathways. Here we describe the first example of neurons selective for learned conspecific vocalizations in adult animals--in starlings that have been trained operantly to recognize conspecific songs. The neuronal population is found in a non-primary forebrain auditory region, exhibits increased responses to the set of learned songs compared with novel songs, and shows differential responses to categories of learned songs based on recognition training contingencies. Within the population, many cells respond highly selectively to a subset of specific motifs (acoustic objects) present only in the learned songs. Such neuronal selectivity may contribute to song-recognition behaviour, which in starlings is sensitive to motif identity. In this system, both top-down and bottom-up processes may modify the tuning properties of neurons during recognition learning, giving rise to plastic representations of behaviourally meaningful auditory objects.
Figures
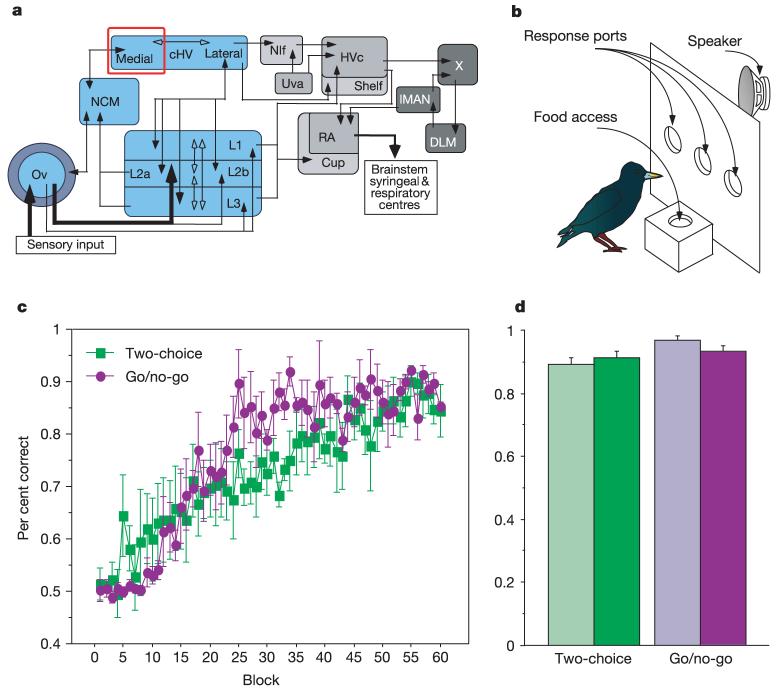
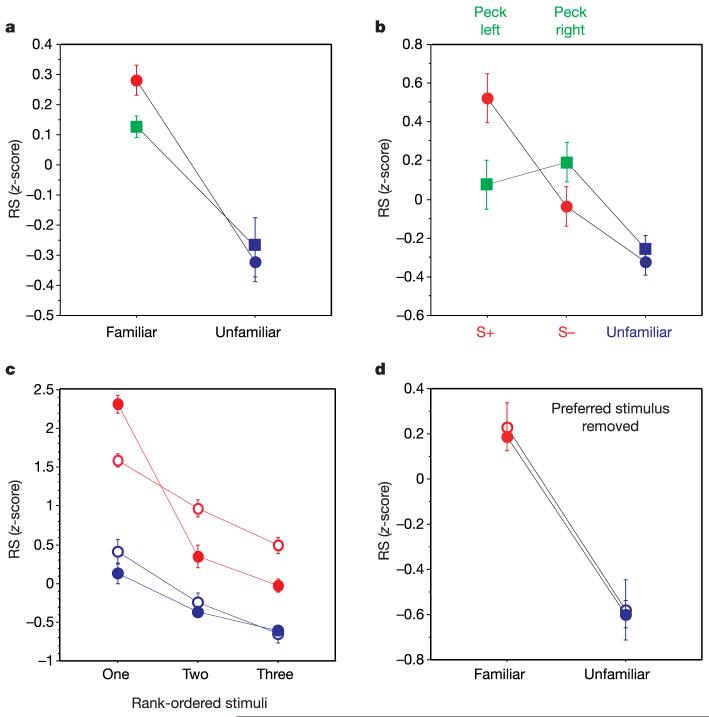
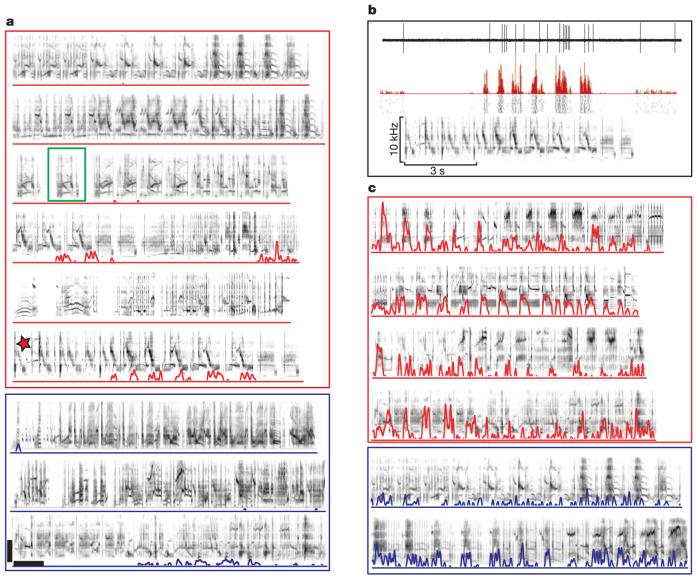
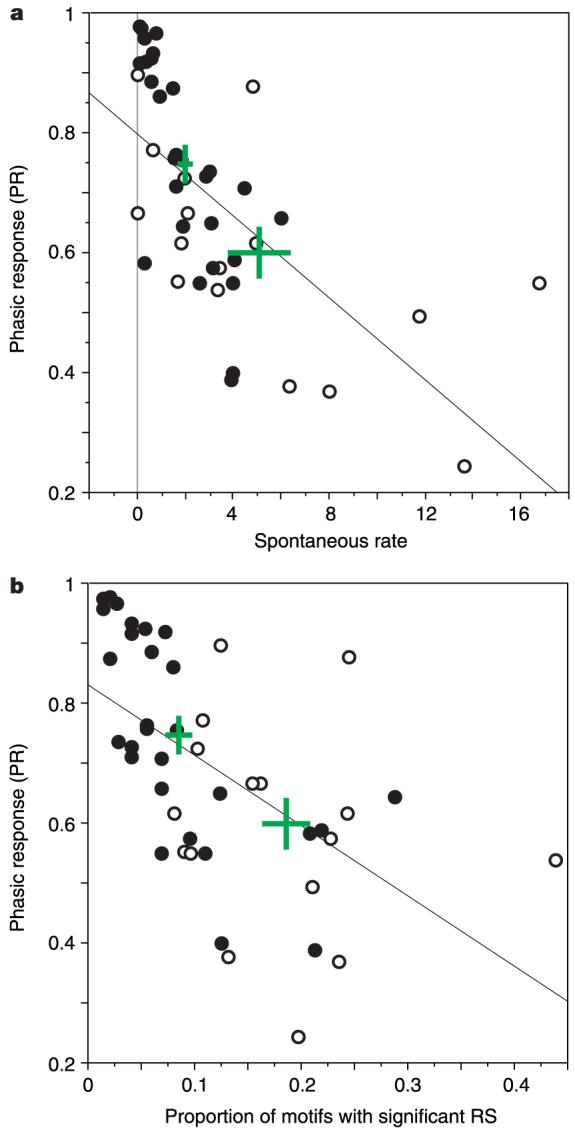
References
-
- Kroodsma DE, Miller EH, editors. The Design of Animal Communication. MIT Press; Cambridge, Massachusetts: 1999.
-
- Gentner TQ, Margoliash D. In: Acoustic Communication. Simmons AM, Popper AN, Fay RR, editors. Springer; New York: 2002. pp. 324–386.
-
- Gentner TQ, Hulse SH, Bentley GE, Ball GF. Individual vocal recognition and the effect of partial lesions to HVc on discrimination, learning, and categorization of conspecific song in adult songbirds. J. Neurobiol. 2000;42:117–133. - PubMed
-
- Gentner TQ, Hulse SH. Perceptual classification based on the component structure of song in European starlings. J. Acoust. Soc. Am. 2000;107:3369–3381. - PubMed
-
- Kroodsma DE, Miller EH, editors. Ecology and Evolution of Acoustic Communication in Birds. Cornell Univ. Press; Ithaca: 1996.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
