

NMR study on the interaction between RPA and DNA decamer containing cis-syn cyclobutane pyrimidine dimer in the presence of XPA: implication for damage verification and strand-specific dual incision in nucleotide excision repair
- PMID: 12907715
- PMCID: PMC169961
- DOI: 10.1093/nar/gkg683
NMR study on the interaction between RPA and DNA decamer containing cis-syn cyclobutane pyrimidine dimer in the presence of XPA: implication for damage verification and strand-specific dual incision in nucleotide excision repair
Abstract
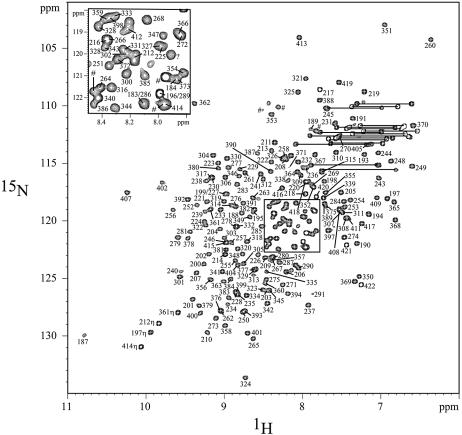
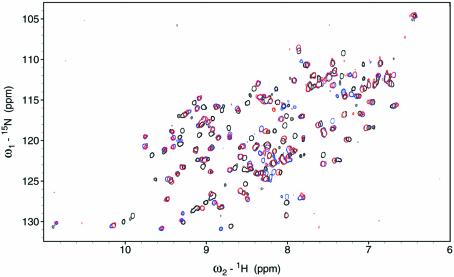
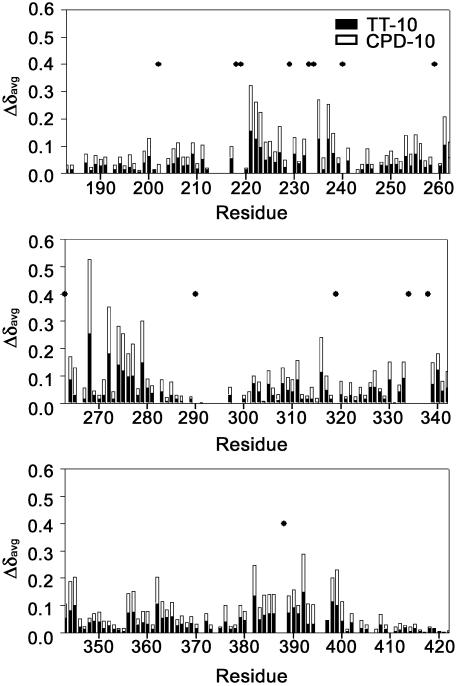
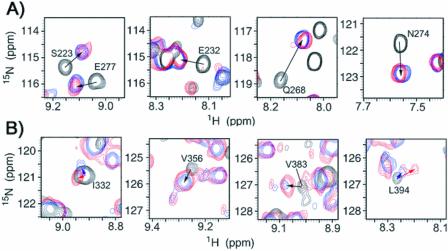
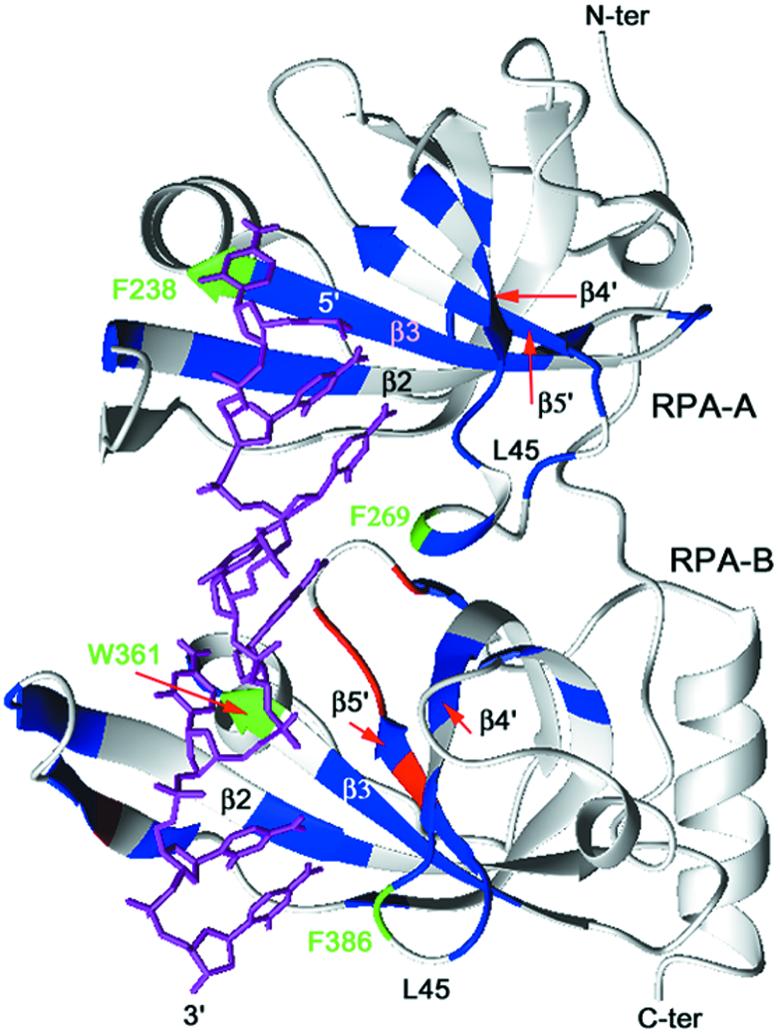
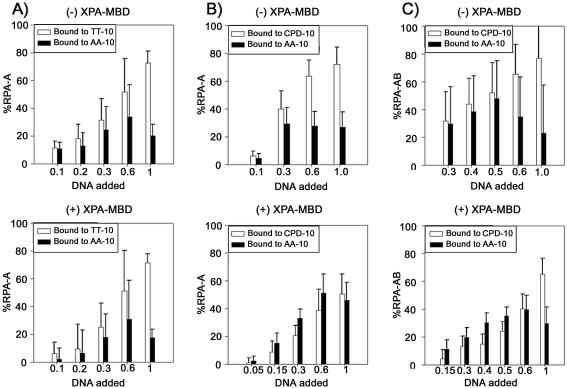
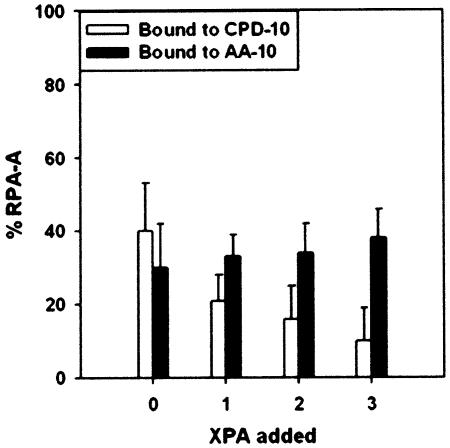
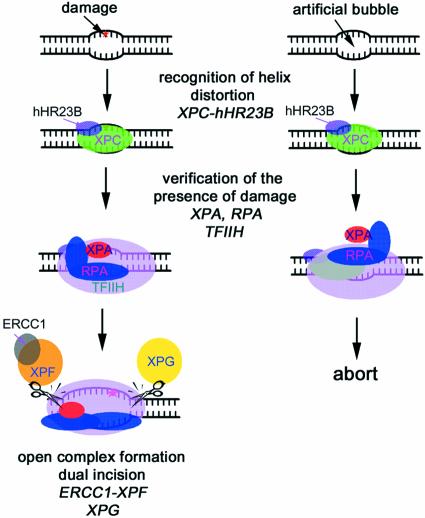
In mammalian cells, nucleotide excision repair (NER) is the major pathway for the removal of bulky DNA adducts. Many of the key NER proteins are members of the XP family (XPA, XPB, etc.), which was named on the basis of its association with the disorder xerodoma pigmentosum. Human replication protein A (RPA), the ubiquitous single-stranded DNA-binding protein, is another of the essential proteins for NER. RPA stimulates the interaction of XPA with damaged DNA by forming an RPA-XPA complex on damaged DNA sites. Binding of RPA to the undamaged DNA strand is most important during NER, because XPA, which directs the excision nucleases XPG and XPF, must bind to the damaged strand. In this study, nuclear magnetic resonance (NMR) spectroscopy was used to assess the binding of the tandem high affinity DNA-binding domains, RPA-AB, and of the isolated domain RPA-A, to normal DNA and damaged DNA containing the cyclobutane pyrimidine dimer (CPD) lesion. Both RPA-A and RPA-AB were found to bind non- specifically to both strands of normal and CPD- containing DNA duplexes. There were no differences observed when binding to normal DNA duplex was examined in the presence of the minimal DNA-binding domain of XPA (XPA-MBD). However, there is a drastic difference for CPD-damaged DNA duplex as both RPA-A and RPA-AB bind specifically to the undamaged strand. The strand-specific binding of RPA and XPA to the damaged duplex DNA shows that RPA and XPA play crucial roles in damage verification and guiding cleavage of damaged DNA during NER.
Figures
References
-
- Wood R.D. (1999) DNA damage recognition during nucleotide excision repair in mammalian cells. Biochimie, 81, 39–44. - PubMed
-
- deLaat W.L., Jaspers,N.G.J. and Hoeijmakers,J.H. (1999) Molecular mechanism of nucleotide excision repair. Genes Dev., 13, 768–785. - PubMed
-
- Batty D.P. and Wood,R.D. (2000) Damage recognition in nucleotide excision repair of DNA. Gene, 241, 193–204. - PubMed
-
- Jones C.J. and Wood,R.D. (1993) Preferential binding of the xeroderma pigmentosum group A complementing protein to damaged DNA. Biochemistry, 32, 12096–12104. - PubMed
-
- He Z., Henricksen,L.A., Wold,M.S. and Ingles,C.J. (1995) RPA involvement in the damage-recognition and incision steps of nucleotide excision repair. Nature, 374, 566–569. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
