

Zinc finger-dependent HIV-1 nucleocapsid protein-TAR RNA interactions
- PMID: 12907727
- PMCID: PMC169955
- DOI: 10.1093/nar/gkg679
Zinc finger-dependent HIV-1 nucleocapsid protein-TAR RNA interactions
Abstract
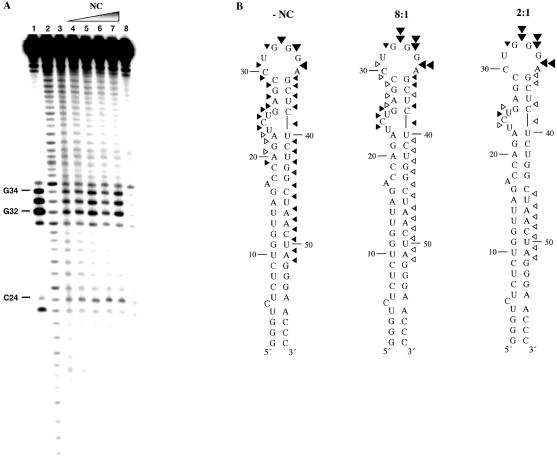
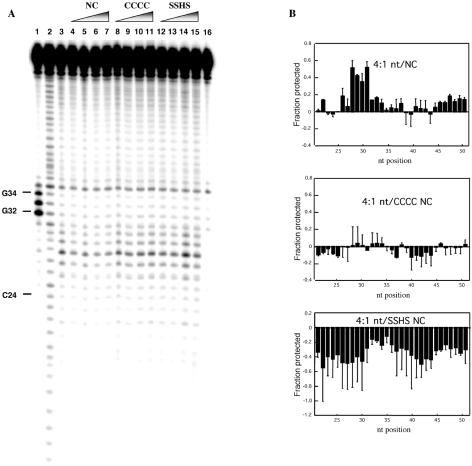
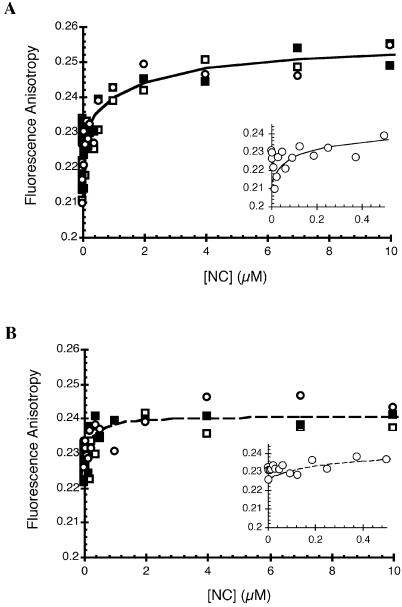
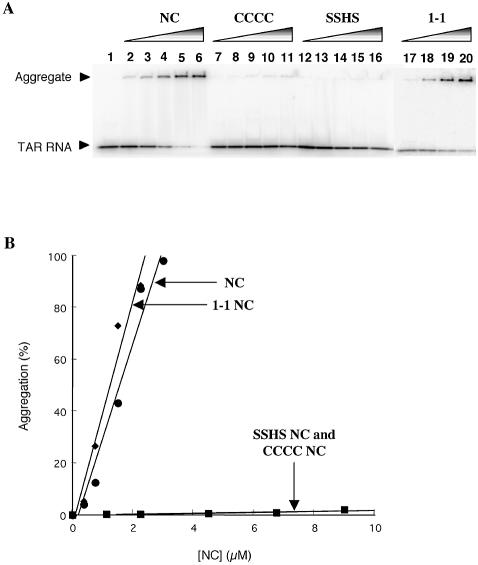
In the minus-strand transfer step of HIV-1 reverse transcription, the nucleocapsid protein (NC) promotes annealing of the 3' 'R' (repeat) region of the RNA genome to its complementary sequence located in the newly synthesized minus-strand strong-stop DNA. The R region contains the highly stable transactivation response (TAR) RNA hairpin. To gain insights into the molecular details of TAR RNA-NC interactions, we carried out hydroxyl radical footprinting, as well as gel-shift and fluorescence anisotropy binding assays using wild-type and mutant forms of NC. Our results support the conclusion that NC variants with mutations in their zinc finger domains have dramatically altered TAR RNA binding interactions relative to wild-type NC. These data demonstrate that a specific zinc finger architecture is required for optimal TAR RNA binding, and help to explain the requirement for the zinc finger motifs of NC in its role as a nucleic acid chaperone in minus-strand transfer.
Figures
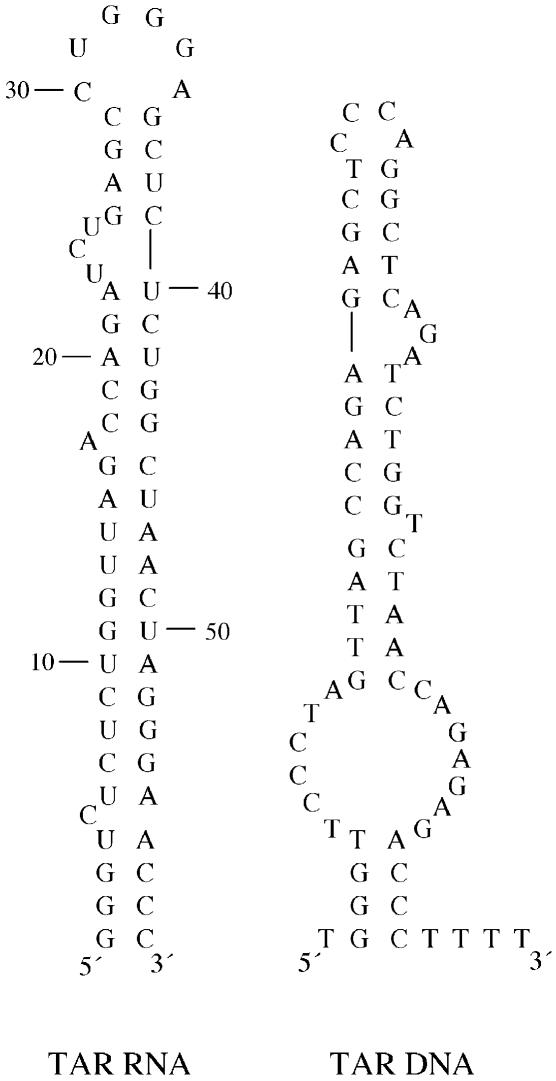

Similar articles
-
Nucleic acid conformational changes essential for HIV-1 nucleocapsid protein-mediated inhibition of self-priming in minus-strand transfer.J Mol Biol. 2003 Jan 3;325(1):1-10. doi: 10.1016/s0022-2836(02)01177-4. J Mol Biol. 2003. PMID: 12473448
-
Mechanistic studies of mini-TAR RNA/DNA annealing in the absence and presence of HIV-1 nucleocapsid protein.J Mol Biol. 2006 Oct 13;363(1):244-61. doi: 10.1016/j.jmb.2006.08.039. Epub 2006 Aug 22. J Mol Biol. 2006. PMID: 16962137
-
Specific interactions between HIV-1 nucleocapsid protein and the TAR element.J Mol Biol. 2005 May 20;348(5):1059-77. doi: 10.1016/j.jmb.2005.03.046. Epub 2005 Apr 7. J Mol Biol. 2005. PMID: 15854644
-
Nucleic acid chaperone activity of HIV-1 nucleocapsid protein: critical role in reverse transcription and molecular mechanism.Prog Nucleic Acid Res Mol Biol. 2005;80:217-86. doi: 10.1016/S0079-6603(05)80006-6. Prog Nucleic Acid Res Mol Biol. 2005. PMID: 16164976 Review. No abstract available.
-
Nucleocapsid protein function in early infection processes.Virus Res. 2008 Jun;134(1-2):39-63. doi: 10.1016/j.virusres.2007.12.006. Epub 2008 Feb 14. Virus Res. 2008. PMID: 18279991 Free PMC article. Review.
Cited by
-
Arterivirus subgenomic mRNA synthesis and virion biogenesis depend on the multifunctional nsp1 autoprotease.J Virol. 2007 Oct;81(19):10496-505. doi: 10.1128/JVI.00683-07. Epub 2007 Jul 11. J Virol. 2007. PMID: 17626105 Free PMC article.
-
Interaction of viral proteins with metal ions: role in maintaining the structure and functions of viruses.FEMS Immunol Med Microbiol. 2005 Feb 1;43(2):105-14. doi: 10.1016/j.femsim.2004.11.004. FEMS Immunol Med Microbiol. 2005. PMID: 15681139 Free PMC article. Review.
-
Fidelity of plus-strand priming requires the nucleic acid chaperone activity of HIV-1 nucleocapsid protein.Nucleic Acids Res. 2009 Apr;37(6):1755-66. doi: 10.1093/nar/gkn1045. Epub 2009 Jan 21. Nucleic Acids Res. 2009. PMID: 19158189 Free PMC article.
-
Role of Divalent Cations in HIV-1 Replication and Pathogenicity.Viruses. 2020 Apr 21;12(4):471. doi: 10.3390/v12040471. Viruses. 2020. PMID: 32326317 Free PMC article. Review.
-
Deaminase-independent inhibition of HIV-1 reverse transcription by APOBEC3G.Nucleic Acids Res. 2007;35(21):7096-108. doi: 10.1093/nar/gkm750. Epub 2007 Oct 16. Nucleic Acids Res. 2007. PMID: 17942420 Free PMC article.
References
-
- South T.L., Kim,B., Hare,D.R. and Summers,M.F. (1990) Zinc fingers and molecular recognition: structure and nucleic acid binding studies of an HIV zinc finger-like domain. Biochem. Pharmacol., 40, 123–129. - PubMed
-
- Summers M.F., Henderson,L.E., Chance,M.R., Bess,J.W.,Jr, South,T.L., Blake,P. R., Sagi,I., Perez-Alvarado,G., Sowder,R.C.,III, Hare,D.R. et al. (1992) Nucleocapsid zinc fingers detected in retroviruses: EXAFS studies of intact viruses and the solution-state structure of the nucleocapsid protein from HIV-1. Protein Sci., 1, 563–574. - PMC - PubMed
-
- Berg J.M. (1986) Potential metal-binding domains in nucleic acid binding proteins. Science, 232, 485–487. - PubMed
-
- Darlix J.L., Lapadat-Tapolsky,M., de Rocquigny,H. and Roques,B.P. (1995) First glimpses at structure-function relationships of the nucleocapsid protein of retroviruses. J. Mol. Biol., 254, 523–537. - PubMed
