

Long-distance root-to-shoot transport of phytochelatins and cadmium in Arabidopsis
- PMID: 12909714
- PMCID: PMC187785
- DOI: 10.1073/pnas.1734072100
Long-distance root-to-shoot transport of phytochelatins and cadmium in Arabidopsis
Abstract
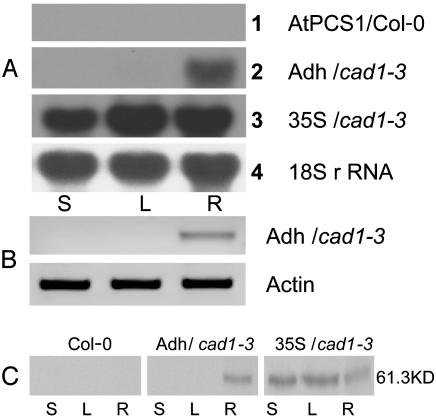
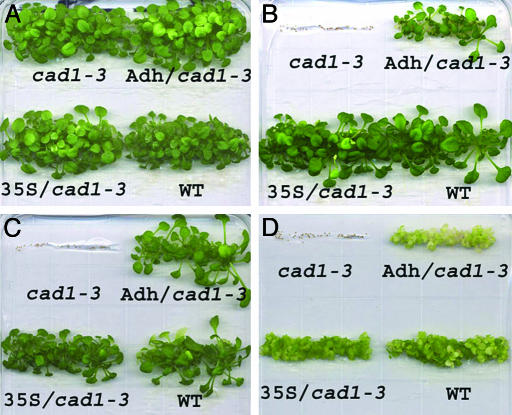
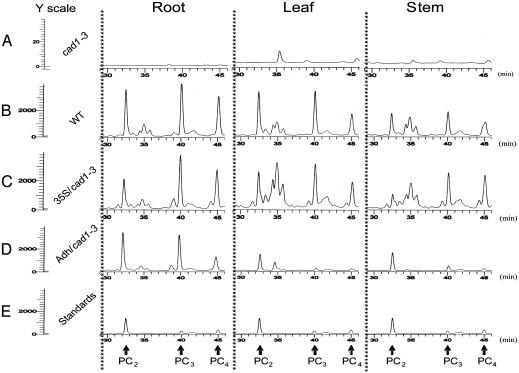
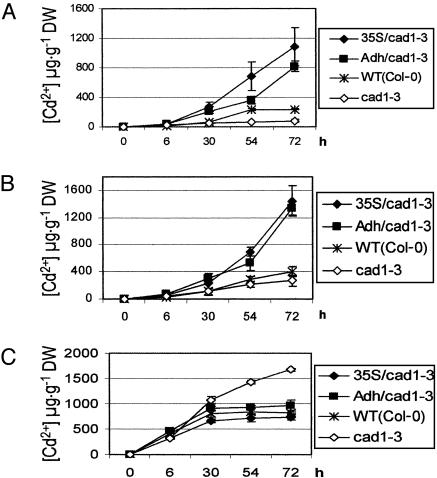
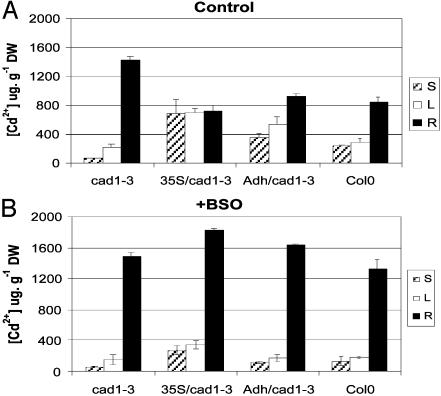
Phytochelatin synthases (PCS) mediate cellular heavy-metal resistance in plants, fungi, and worms. However, phytochelatins (PCs) are generally considered to function as intracellular heavy-metal detoxification mechanisms, and whether long-distance transport of PCs occurs during heavy-metal detoxification remains unknown. Here, wheat TaPCS1 cDNA expression was either targeted to Arabidopsis roots with the Arabidopsis alcohol dehydrogenase (Adh) promoter (Adh::TaPCS1/cad1-3) or ectopically expressed with the cauliflower mosaic virus 35S promoter (35S::TaPCS1/cad1-3) in the PC-deficient mutant cad1-3. Adh::TaPCS1/cad1-3 and 35S::TaPCS1/cad1-3 complemented the cadmium, mercury, and arsenic sensitivities of the cad1-3 mutant. Northern blot, RT-PCR, and Western blot analyses showed Adh promoter-driven TaPCS1 expression only in roots and thus demonstrated lack of long-distance TaPCS1 mRNA and protein transport in plants. Fluorescence HPLC analyses showed that under Cd2+ stress, no PCs were detectable in cad1-3. However, in Adh::TaPCS1/cad1-3 plants, PCs were detected in roots and in rosette leaves and stems. Inductively coupled plasma atomic emission spectrometer analyses showed that either root-specific or ectopic expression of TaPCS1 significantly enhanced long-distance Cd2+ transport into stems and rosette leaves. Unexpectedly, transgenic expression of TaPCS1 reduced Cd2+ accumulation in roots compared with cad1-3. The reduced Cd2+ accumulation in roots and enhanced root-to-shoot Cd2+ transport in transgenic plants were abrogated by l-buthionine sulfoximine. The presented findings show that (i) transgenic expression of TaPCS1 suppresses the heavy-metal sensitivity of cad1-3, (ii) PCs can be transported from roots to shoots, and (iii) transgenic expression of the TaPCS1 gene increases long-distance root-to-shoot Cd2+ transport and reduces Cd2+ accumulation in roots.
Figures

Similar articles
-
An improved grafting technique for mature Arabidopsis plants demonstrates long-distance shoot-to-root transport of phytochelatins in Arabidopsis.Plant Physiol. 2006 May;141(1):108-20. doi: 10.1104/pp.105.072637. Epub 2006 Mar 10. Plant Physiol. 2006. PMID: 16531489 Free PMC article.
-
Fission yeast HMT1 lowers seed cadmium through phytochelatin-dependent vacuolar sequestration in Arabidopsis.Plant Physiol. 2012 Apr;158(4):1779-88. doi: 10.1104/pp.111.192872. Epub 2012 Feb 7. Plant Physiol. 2012. PMID: 22319073 Free PMC article.
-
Tolerance to toxic metals by a gene family of phytochelatin synthases from plants and yeast.EMBO J. 1999 Jun 15;18(12):3325-33. doi: 10.1093/emboj/18.12.3325. EMBO J. 1999. PMID: 10369673 Free PMC article.
-
Phytochelatins and their roles in heavy metal detoxification.Plant Physiol. 2000 Jul;123(3):825-32. doi: 10.1104/pp.123.3.825. Plant Physiol. 2000. PMID: 10889232 Free PMC article. Review. No abstract available.
-
Phytochelatins: Sulfur-Containing Metal(loid)-Chelating Ligands in Plants.Int J Mol Sci. 2023 Jan 26;24(3):2430. doi: 10.3390/ijms24032430. Int J Mol Sci. 2023. PMID: 36768751 Free PMC article. Review.
Cited by
-
Characterization of Brassica rapa metallothionein and phytochelatin synthase genes potentially involved in heavy metal detoxification.PLoS One. 2021 Jun 4;16(6):e0252899. doi: 10.1371/journal.pone.0252899. eCollection 2021. PLoS One. 2021. PMID: 34086824 Free PMC article.
-
Exogenous abscisic acid application decreases cadmium accumulation in Arabidopsis plants, which is associated with the inhibition of IRT1-mediated cadmium uptake.Front Plant Sci. 2014 Dec 16;5:721. doi: 10.3389/fpls.2014.00721. eCollection 2014. Front Plant Sci. 2014. PMID: 25566293 Free PMC article.
-
Loss of function of Arabidopsis C-terminal domain phosphatase-like1 activates iron deficiency responses at the transcriptional level.Plant Physiol. 2013 Jan;161(1):330-45. doi: 10.1104/pp.112.207043. Epub 2012 Nov 9. Plant Physiol. 2013. PMID: 23144187 Free PMC article.
-
Overexpression of Arabidopsis phytochelatin synthase in tobacco plants enhances Cd(2+) tolerance and accumulation but not translocation to the shoot.Planta. 2006 Jan;223(2):180-90. doi: 10.1007/s00425-005-0073-3. Epub 2005 Aug 20. Planta. 2006. PMID: 16133212
-
Metal species involved in long distance metal transport in plants.Front Plant Sci. 2014 Mar 25;5:105. doi: 10.3389/fpls.2014.00105. eCollection 2014. Front Plant Sci. 2014. PMID: 24723928 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
