

Spatial restriction of alpha4 integrin phosphorylation regulates lamellipodial stability and alpha4beta1-dependent cell migration
- PMID: 12913113
- PMCID: PMC2173787
- DOI: 10.1083/jcb.200304031
Spatial restriction of alpha4 integrin phosphorylation regulates lamellipodial stability and alpha4beta1-dependent cell migration
Abstract
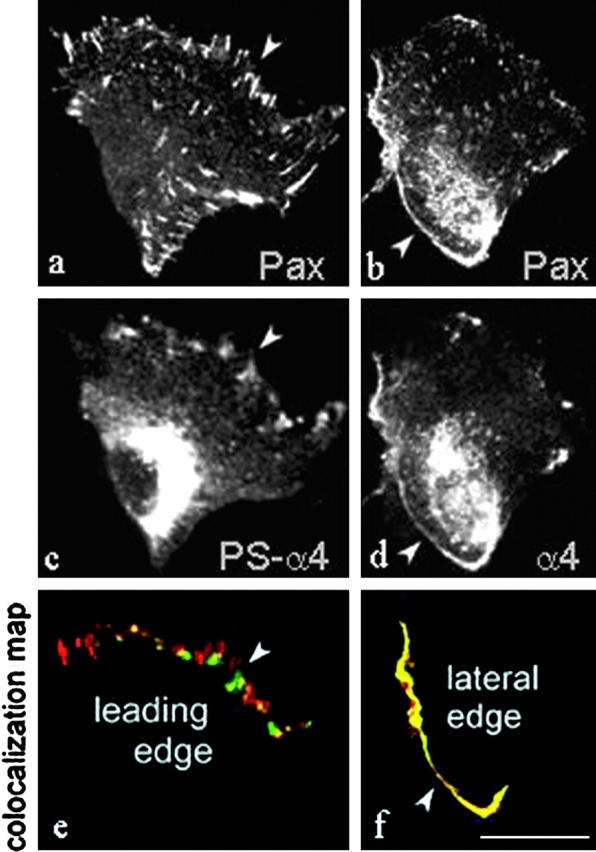
Integrins coordinate spatial signaling events essential for cell polarity and directed migration. Such signals from alpha4 integrins regulate cell migration in development and in leukocyte trafficking. Here, we report that efficient alpha4-mediated migration requires spatial control of alpha4 phosphorylation by protein kinase A, and hence localized inhibition of binding of the signaling adaptor, paxillin, to the integrin. In migrating cells, phosphorylated alpha4 accumulated along the leading edge. Blocking alpha4 phosphorylation by mutagenesis or by inhibition of protein kinase A drastically reduced alpha4-dependent migration and lamellipodial stability. alpha4 phosphorylation blocks paxillin binding in vitro; we now find that paxillin and phospho-alpha4 were in distinct clusters at the leading edge of migrating cells, whereas unphosphorylated alpha4 and paxillin colocalized along the lateral edges of those cells. Furthermore, enforced paxillin association with alpha4 inhibits migration and reduced lamellipodial stability. These results show that topographically specific integrin phosphorylation can control cell migration and polarization by spatial segregation of adaptor protein binding.
Figures










Similar articles
-
An alpha4 integrin-paxillin-Arf-GAP complex restricts Rac activation to the leading edge of migrating cells.Nat Cell Biol. 2005 Apr;7(4):343-52. doi: 10.1038/ncb1234. Epub 2005 Mar 27. Nat Cell Biol. 2005. PMID: 15793570
-
Binding of paxillin to alpha4 integrins modifies integrin-dependent biological responses.Nature. 1999 Dec 9;402(6762):676-81. doi: 10.1038/45264. Nature. 1999. PMID: 10604475
-
The role of the alpha4 integrin-paxillin interaction in regulating leukocyte trafficking.Exp Mol Med. 2006 Jun 30;38(3):191-5. doi: 10.1038/emm.2006.23. Exp Mol Med. 2006. PMID: 16819276 Review.
-
An integrin-alpha4-14-3-3zeta-paxillin ternary complex mediates localised Cdc42 activity and accelerates cell migration.J Cell Sci. 2009 May 15;122(Pt 10):1654-64. doi: 10.1242/jcs.049130. Epub 2009 Apr 28. J Cell Sci. 2009. PMID: 19401330 Free PMC article.
-
New approaches to blockade of alpha4-integrins, proven therapeutic targets in chronic inflammation.Biochem Pharmacol. 2006 Nov 30;72(11):1460-8. doi: 10.1016/j.bcp.2006.06.014. Epub 2006 Jul 25. Biochem Pharmacol. 2006. PMID: 16870156 Review.
Cited by
-
Hyperglycemia reduces integrin subunits alpha v and alpha 5 on the surface of dermal fibroblasts contributing to deficient migration.Mol Cell Biochem. 2016 Oct;421(1-2):19-28. doi: 10.1007/s11010-016-2780-4. Epub 2016 Aug 18. Mol Cell Biochem. 2016. PMID: 27535240
-
Neurite outgrowth on a fibronectin isoform expressed during peripheral nerve regeneration is mediated by the interaction of paxillin with alpha4beta1 integrins.BMC Neurosci. 2007 Jun 29;8:44. doi: 10.1186/1471-2202-8-44. BMC Neurosci. 2007. PMID: 17603879 Free PMC article.
-
HER2 carboxyl-terminal fragments regulate cell migration and cortactin phosphorylation.J Biol Chem. 2009 Sep 11;284(37):25302-13. doi: 10.1074/jbc.M109.001982. Epub 2009 Jul 8. J Biol Chem. 2009. PMID: 19589785 Free PMC article.
-
The PIX-GIT complex: a G protein signaling cassette in control of cell shape.Semin Cell Dev Biol. 2008 Jun;19(3):234-44. doi: 10.1016/j.semcdb.2008.01.002. Epub 2008 Jan 20. Semin Cell Dev Biol. 2008. PMID: 18299239 Free PMC article. Review.
-
Roles of Integrins in Gastrointestinal Cancer Metastasis.Front Mol Biosci. 2021 Nov 15;8:708779. doi: 10.3389/fmolb.2021.708779. eCollection 2021. Front Mol Biosci. 2021. PMID: 34869579 Free PMC article. Review.
References
-
- Alt, A., M. Ohba, L. Li, M. Gartsbein, A. Belanger, M.F. Denning, T. Kuroki, S.H. Yuspa, and T. Tennenbaum. 2001. Protein kinase Cdelta-mediated phosphorylation of alpha6beta4 is associated with reduced integrin localization to the hemidesmosome and decreased keratinocyte attachment. Cancer Res. 61:4591–4598. - PubMed
-
- Arroyo, A.G., J.T. Yang, H. Rayburn, and R.O. Hynes. 1996. Differential requirements for α4 integrins in hematopoiesis. Cell. 85:997–1008. - PubMed
-
- Baker, S.E., O. Skalli, R.D. Goldman, and J.C. Jones. 1997. Laminin-5 and modulation of keratin cytoskeleton arrangement in FG pancreatic carcinoma cells: involvement of IFAP300 and evidence that laminin-5/cell interactions correlate with a dephosphorylation of alpha 6A integrin. Cell Motil. Cytoskeleton. 37:271–286. - PubMed
-
- Calderwood, D.A., R. Zent, R. Grant, D.J. Rees, R.O. Hynes, and M.H. Ginsberg. 1999. The Talin head domain binds to integrin beta subunit cytoplasmic tails and regulates integrin activation. J. Biol. Chem. 274:28071–28074. - PubMed
-
- Chan, B.M.C., P.D. Kassner, J.A. Schiro, H.R. Byers, T.S. Kupper, and M.E. Hemler. 1992. Distinct cellular functions mediated by different VLA integrin α subunit cytoplasmic domains. Cell. 68:1051–1060. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
