

PAUSED, a putative exportin-t, acts pleiotropically in Arabidopsis development but is dispensable for viability
- PMID: 12913148
- PMCID: PMC181277
- DOI: 10.1104/pp.103.023291
PAUSED, a putative exportin-t, acts pleiotropically in Arabidopsis development but is dispensable for viability
Abstract
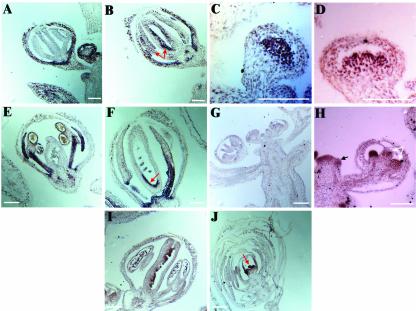
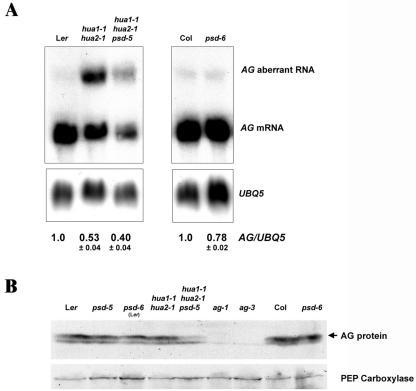
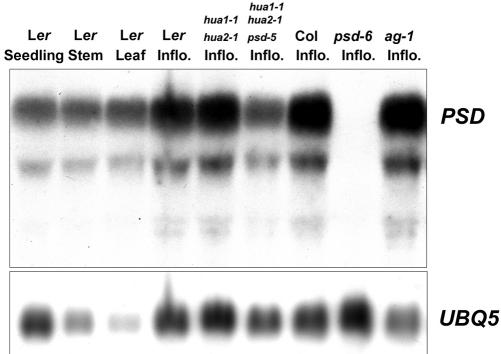
Exportin-t was first identified in humans as a protein that mediates the export of tRNAs from the nucleus to the cytoplasm. Mutations in Los1p, the Saccharomyces cerevisiae exportin-t homolog, result in nuclear accumulation of tRNAs. Because no exportin-t mutants have been reported in multicellular organisms, the developmental functions of exportin-t have not been determined. Here, we report the isolation and characterization of two Arabidopsis exportin-t mutants, paused-5 and paused-6. The mutant phenotypes indicate that exportin-t acts pleiotropically in plant development. In particular, paused-5 and paused-6 result in delayed leaf formation during vegetative development. The two paused mutations also cause the transformation of reproductive organs into perianth organs in the hua1-1 hua2-1 background, which is partially defective in reproductive organ identity specification. The floral phenotypes of hua1-1 hua2-1 paused mutants resemble those of mutations in the floral homeotic gene AGAMOUS. Moreover, paused-5 enhances the mutant phenotypes of two floral meristem identity genes, LEAFY and APETALA1. The developmental defects caused by paused mutations confirm the important roles of exportin-t in gene expression in multicellular organisms. In addition, a paused null allele, paused-6, is still viable, suggesting the presence of redundant tRNA export pathway(s) in Arabidopsis.
Figures




References
-
- Arts GJ, Fornerod M, Mattaj IW (1998) Identification of a nuclear export receptor for tRNA. Curr Biol 8: 305–314 - PubMed
-
- Bollman KM, Aukerman MJ, Park M-Y, Hunter C, Berardini TZ, Poethig RS (2003) HASTY, the Arabidopsis ortholog of exportin 5/MSN5, regulates phase change and morphogenesis. Development 130: 1493–1504 - PubMed
-
- Bowman JL, Alvarez J, Weigel D, Meyerowitz EM, Smyth DR (1993) Control of flower development in Arabidopsis thaliana by APETALA1 and interacting genes. Development 119: 721–743
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
