

BWMK1, a rice mitogen-activated protein kinase, locates in the nucleus and mediates pathogenesis-related gene expression by activation of a transcription factor
- PMID: 12913152
- PMCID: PMC181281
- DOI: 10.1104/pp.103.023176
BWMK1, a rice mitogen-activated protein kinase, locates in the nucleus and mediates pathogenesis-related gene expression by activation of a transcription factor
Abstract
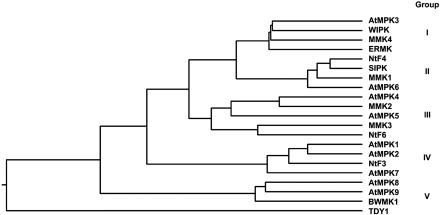
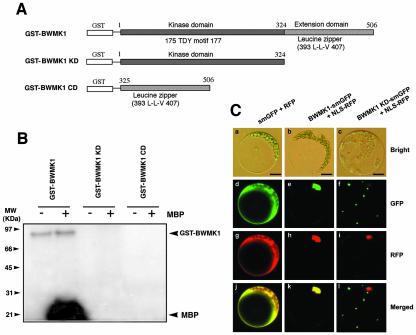
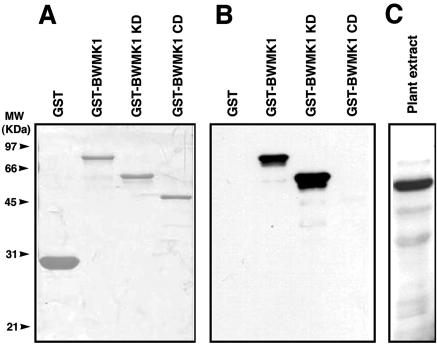
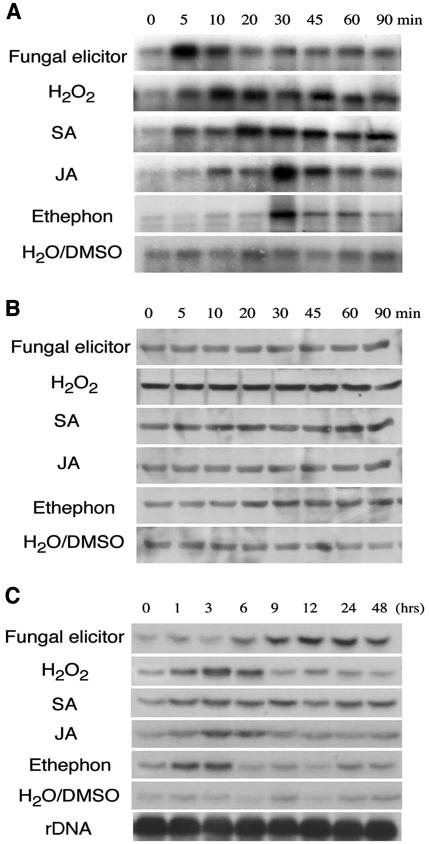
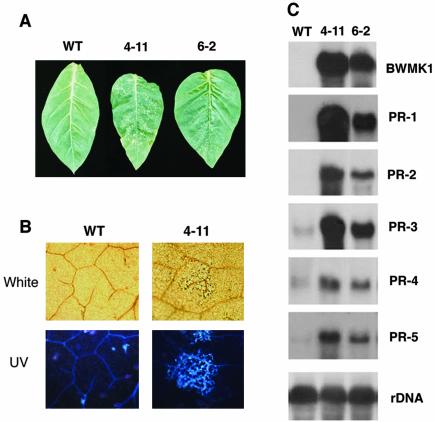
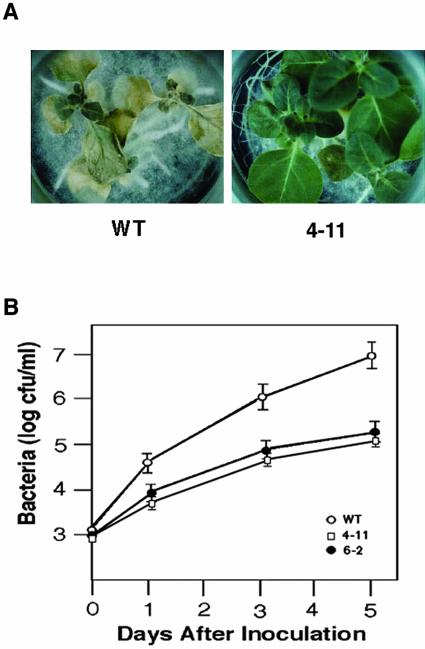
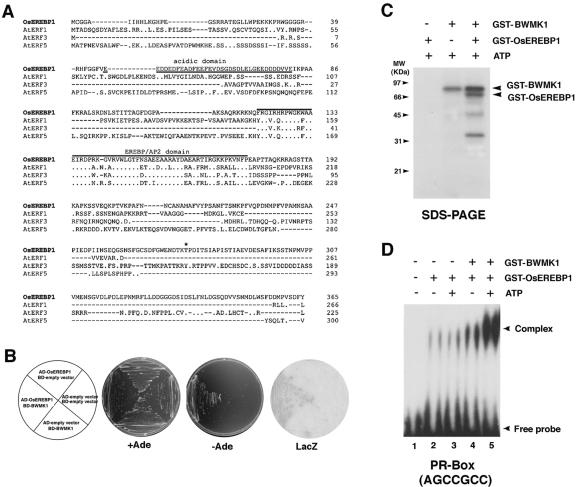
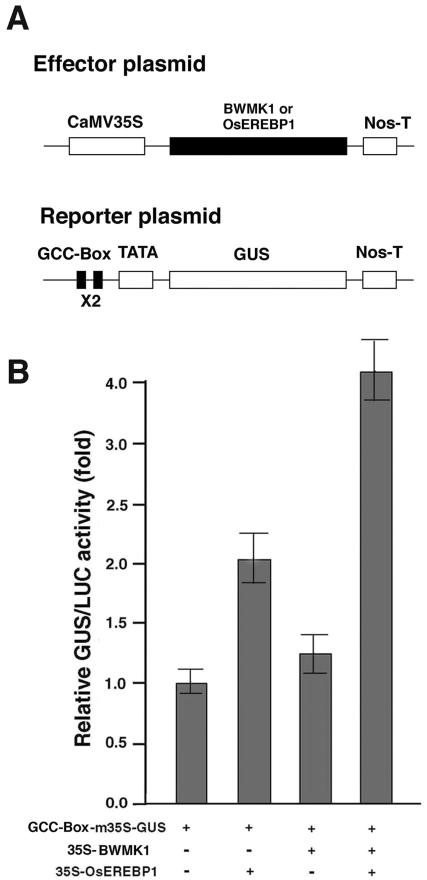
Mitogen-activated protein kinase (MAPK) cascades are known to transduce plant defense signals, but the downstream components of the MAPK have as yet not been elucidated. Here, we report an MAPK from rice (Oryza sativa), BWMK1, and a transcription factor, OsEREBP1, phosphorylated by the kinase. The MAPK carries a TDY phosphorylation motif instead of the more common TEY motif in its kinase domain and has an unusually extended C-terminal domain that is essential to its kinase activity and translocation to the nucleus. The MAPK phosphorylates OsEREBP1 that binds to the GCC box element (AGCCGCC) of the several basic pathogenesis-related gene promoters, which in turn enhances DNA-binding activity of the factor to the cis element in vitro. Transient co-expression of the BWMK1 and OsEREBP1 in Arabidopsis protoplasts elevates the expression of the beta-glucuronidase reporter gene driven by the GCC box element. Furthermore, transgenic tobacco (Nicotiana tabacum) plants overexpressing BWMK1 expressed many pathogenesis-related genes at higher levels than wild-type plants with an enhanced resistance to pathogens. These findings suggest that MAPKs contribute to plant defense signal transduction by phosphorylating one or more transcription factors.
Figures
References
-
- Abel S, Theologis A (1994) Transient transformation of Arabidopsis leaf protoplasts: a versatile experimental system to study gene expression. Plant J 5: 421–427 - PubMed
-
- Agrawal GK, Rakwal R, Iwahashi H (2002) Isolation of novel rice (Oryza sativa L.) multiple stress responsive MAP kinase gene, OsMSRMK2, whose mRNA accumulates rapidly in response to environmental cues. Biochem Biophys Res Commun 294: 1009–1016 - PubMed
-
- An G, Prebert AM, Ha SB (1988) Binary vectors. In SB Gelvin, RA Schilperoort, eds, Plant Molecular Biology Manual. Academic Press, Dordrecht, The Netherlands, pp 1–19
-
- Asai T, Tena G, Plotnikova J, Willmann MR, Chiu WL, Gomez-Gomez L, Boller T, Ausubel FM, Sheen J (2002) MAP kinase signalling cascade in Arabidopsis innate immunity. Nature 415: 977–983 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
