

A bypass of sucrose synthase leads to low internal oxygen and impaired metabolic performance in growing potato tubers
- PMID: 12913161
- PMCID: PMC181290
- DOI: 10.1104/pp.103.022236
A bypass of sucrose synthase leads to low internal oxygen and impaired metabolic performance in growing potato tubers
Abstract
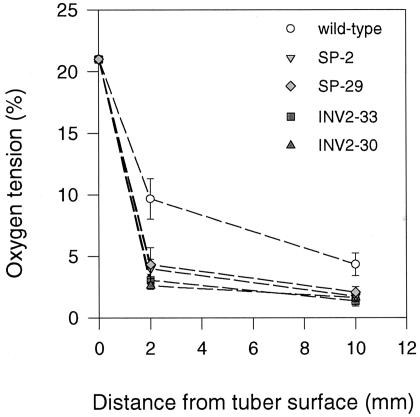
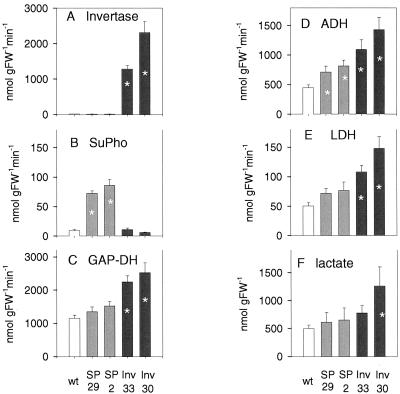
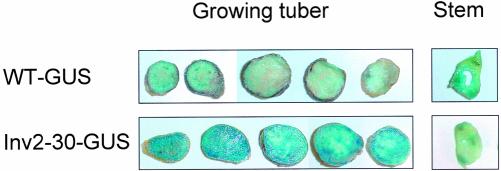
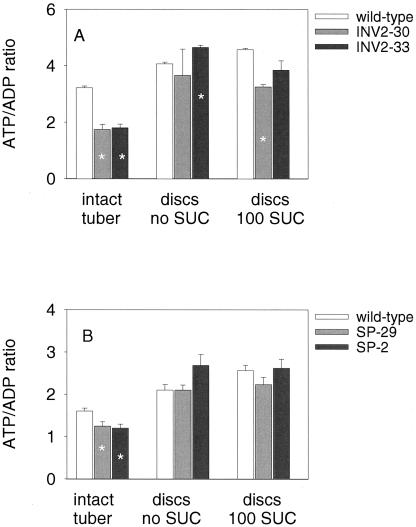
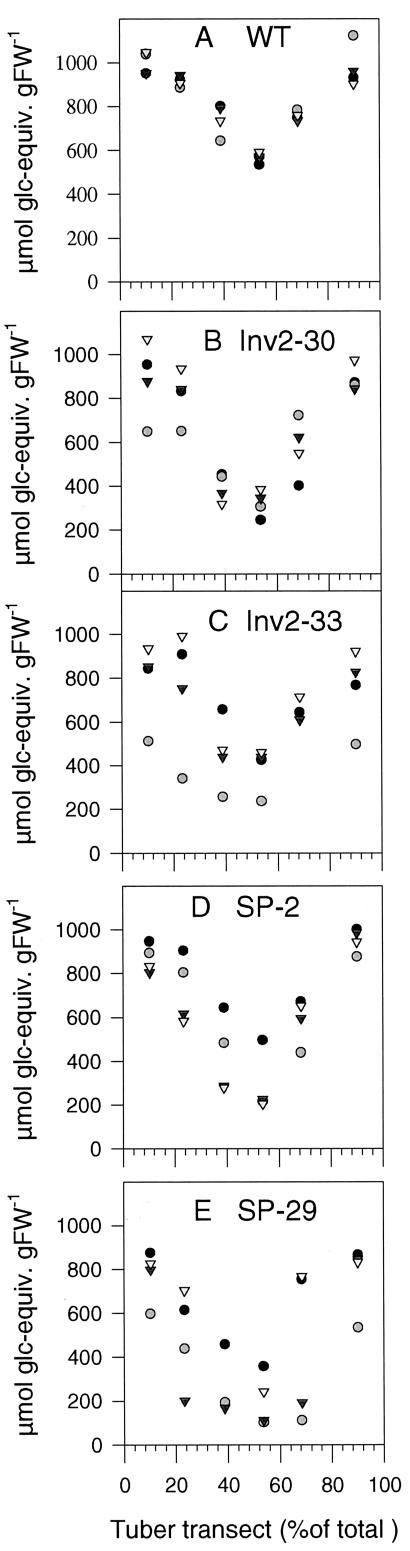
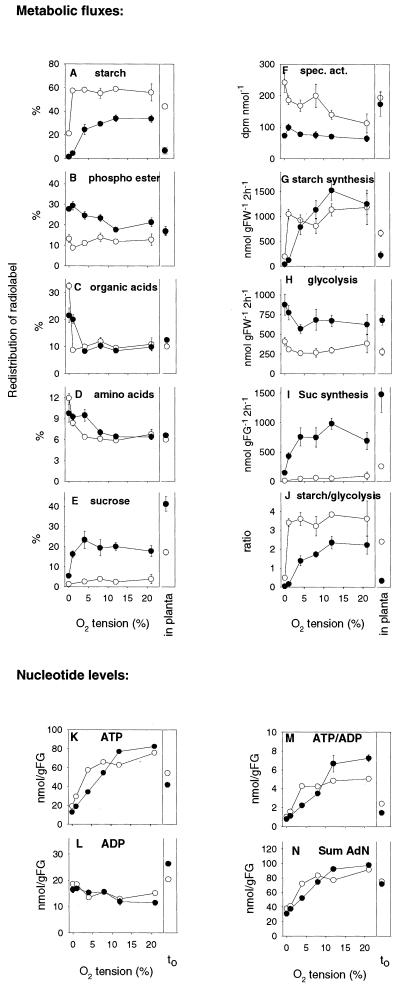
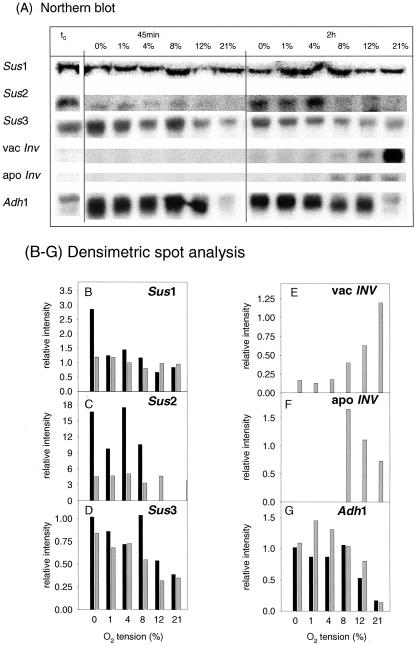
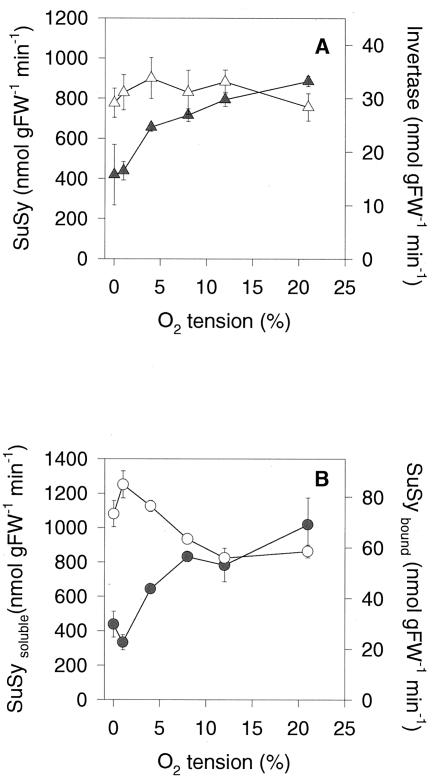
Plants possess two alternative biochemical pathways for sucrose (Suc) degradation. One involves hydrolysis by invertase followed by phosphorylation via hexokinase and fructokinase, and the other route-which is unique to plants-involves a UDP-dependent cleavage of Suc that is catalyzed by Suc synthase (SuSy). In the present work, we tested directly whether a bypass of the endogenous SuSy route by ectopic overexpression of invertase or Suc phosphorylase affects internal oxygen levels in growing tubers and whether this is responsible for their decreased starch content. (a) Oxygen tensions were lower within transgenic tubers than in wild-type tubers. Oxygen tensions decreased within the first 10 mm of tuber tissue, and this gradient was steeper in transgenic tubers. (b) Invertase-overexpressing tubers had higher activities of glyceraldehyde-3-phosphate dehydrogenase, lactate dehydrogenase, and alcohol dehydrogenase, and (c) higher levels of lactate. (d) Expression of a low-oxygen-sensitive Adh1-beta-glucuronidase reporter gene construct was more strongly induced in the invertase-overexpressing background compared with wild-type background. (e) Intact transgenic tubers had lower ATP to ADP ratios than the wild type. ATP to ADP ratio was restored to wild type, when discs of transgenic tubers were incubated at 21% (v/v) oxygen. (f) Starch decreased from the periphery to the center of the tuber. This decrease was much steeper in the transgenic lines, leading to lower starch content especially near the center of the tuber. (g) Metabolic fluxes (based on redistribution of (14)C-glucose) and ATP to ADP ratios were analyzed in more detail, comparing discs incubated at various external oxygen tensions (0%, 1%, 4%, 8%, 12%, and 21% [v/v]) with intact tubers. Discs of Suc phosphorylase-expressing lines had similar ATP to ADP ratios and made starch as fast as wild type in high oxygen but had lower ATP to ADP ratios and lower rates of starch synthesis than wild type at low-oxygen tensions typical to those found inside an intact tuber. (h) In discs of wild-type tubers, subambient oxygen concentrations led to a selective increase in the mRNA levels of specific SuSy genes, whereas the mRNA levels of genes encoding vacuolar and apoplastic invertases decreased. (i) These results imply that repression of invertase and mobilization of Suc via the energetically less costly route provided by SuSy is important in growing tubers because it conserves oxygen and allows higher internal oxygen tensions to be maintained than would otherwise be possible.
Figures
References
-
- Appeldoorn NJG, de Bruijn SM, Koot-Gronsveld EAM, Visser RGF, Vreugdenhil D, van der Plas LHW (1997) Developmental changes of enzymes involved in conversion of sucrose to hexose phosphate during early tuberisation of potato. Planta 202: 220-226
-
- ap Rees T (1984) Sucrose metabolism. In DH Lewis, ed, Storage carbohydrates in Vascular Plants. Cambridge University Press, Cambridge, UK pp 53-73
-
- Avigad G (1982) Sucrose and other disaccharides. In TA Loewus, W Tanner, eds, Encyclopedia of Plant Physiology. Springer-Verlag, Heidelberg, pp 217-347
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
