

The Chlamydomonas reinhardtii cia3 mutant lacking a thylakoid lumen-localized carbonic anhydrase is limited by CO2 supply to rubisco and not photosystem II function in vivo
- PMID: 12913181
- PMCID: PMC181310
- DOI: 10.1104/pp.103.023481
The Chlamydomonas reinhardtii cia3 mutant lacking a thylakoid lumen-localized carbonic anhydrase is limited by CO2 supply to rubisco and not photosystem II function in vivo
Abstract
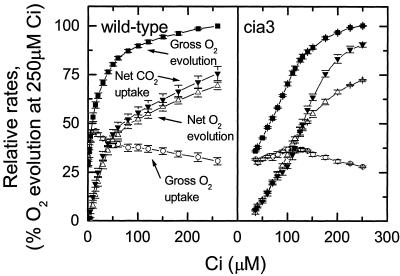
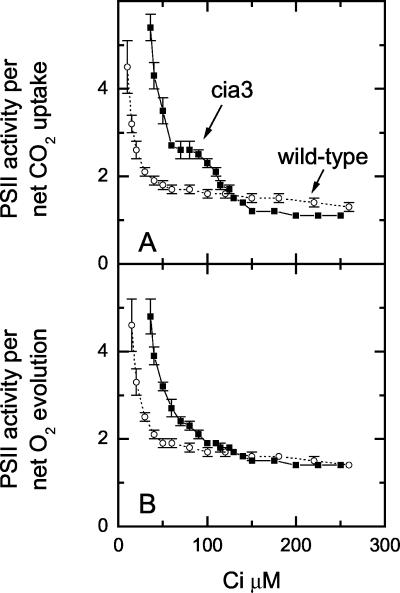
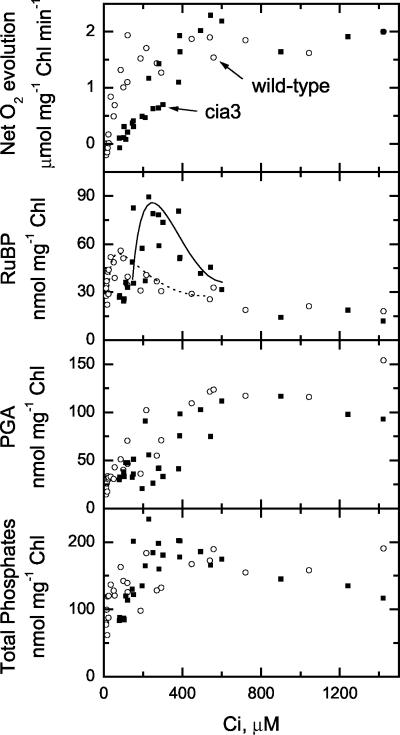
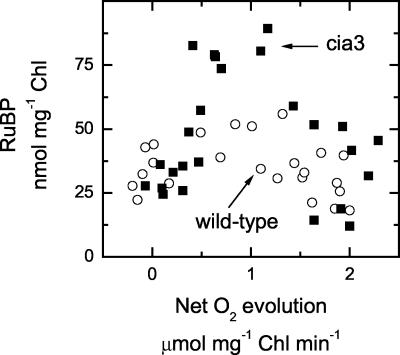
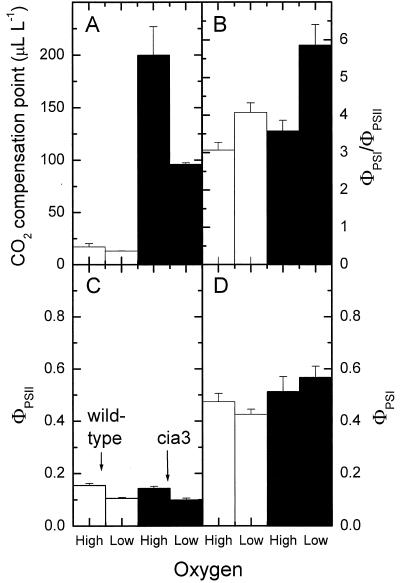
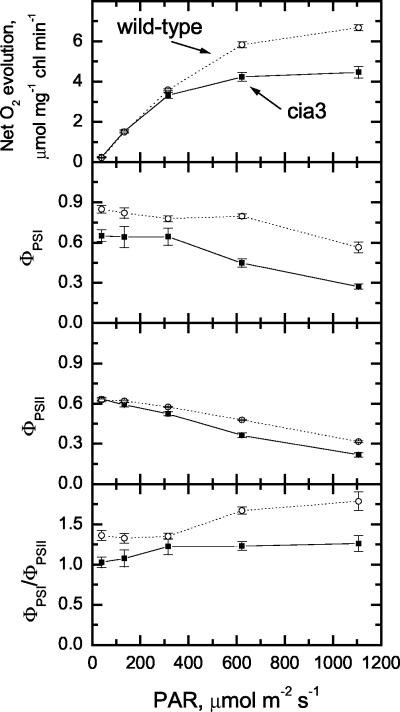
The Chlamydomonas reinhardtii cia3 mutant has a phenotype indicating that it requires high-CO(2) levels for effective photosynthesis and growth. It was initially proposed that this mutant was defective in a carbonic anhydrase (CA) that was a key component of the photosynthetic CO(2)-concentrating mechanism (CCM). However, more recent identification of the genetic lesion as a defect in a lumenal CA associated with photosystem II (PSII) has raised questions about the role of this CA in either the CCM or PSII function. To resolve the role of this lumenal CA, we re-examined the physiology of the cia3 mutant. We confirmed and extended previous gas exchange analyses by using membrane-inlet mass spectrometry to monitor(16)O(2),(18)O(2), and CO(2) fluxes in vivo. The results demonstrate that PSII electron transport is not limited in the cia3 mutant at low inorganic carbon (Ci). We also measured metabolite pools sizes and showed that the RuBP pool does not fall to abnormally low levels at low Ci as might be expected by a photosynthetic electron transport or ATP generation limitation. Overall, the results demonstrate that under low Ci conditions, the mutant lacks the ability to supply Rubisco with adequate CO(2) for effective CO(2) fixation and is not limited directly by any aspect of PSII function. We conclude that the thylakoid CA is primarily required for the proper functioning of the CCM at low Ci by providing an ample supply of CO(2) for Rubisco.
Figures
Similar articles
-
Role of a novel photosystem II-associated carbonic anhydrase in photosynthetic carbon assimilation in Chlamydomonas reinhardtii.FEBS Lett. 1999 Feb 5;444(1):102-5. doi: 10.1016/s0014-5793(99)00037-x. FEBS Lett. 1999. PMID: 10037156
-
Identification and functional role of the carbonic anhydrase Cah3 in thylakoid membranes of pyrenoid of Chlamydomonas reinhardtii.Biochim Biophys Acta. 2012 Aug;1817(8):1248-55. doi: 10.1016/j.bbabio.2012.02.014. Epub 2012 Feb 21. Biochim Biophys Acta. 2012. PMID: 22709623
-
Carbonic anhydrase CAH3 supports the activity of photosystem II under increased pH.Biochim Biophys Acta Bioenerg. 2019 Jul 1;1860(7):582-590. doi: 10.1016/j.bbabio.2019.06.003. Epub 2019 Jun 19. Biochim Biophys Acta Bioenerg. 2019. PMID: 31226314
-
CAH3 from Chlamydomonas reinhardtii: Unique Carbonic Anhydrase of the Thylakoid Lumen.Cells. 2024 Jan 5;13(2):109. doi: 10.3390/cells13020109. Cells. 2024. PMID: 38247801 Free PMC article. Review.
-
The carbonic anhydrase isoforms of Chlamydomonas reinhardtii: intracellular location, expression, and physiological roles.Photosynth Res. 2011 Sep;109(1-3):133-49. doi: 10.1007/s11120-011-9635-3. Epub 2011 Mar 2. Photosynth Res. 2011. PMID: 21365258 Review.
Cited by
-
Fast cadmium inhibition of photosynthesis in cyanobacteria in vivo and in vitro studies using perturbed angular correlation of gamma-rays.J Biol Inorg Chem. 2006 Sep;11(6):725-34. doi: 10.1007/s00775-006-0113-x. Epub 2006 Jul 5. J Biol Inorg Chem. 2006. PMID: 16821039
-
A Rapid Method for Detecting Normal or Modified Plant and Algal Carbonic Anhydrase Activity Using Saccharomyces cerevisiae.Plants (Basel). 2022 Jul 20;11(14):1882. doi: 10.3390/plants11141882. Plants (Basel). 2022. PMID: 35890517 Free PMC article.
-
Control of Rubisco function via homeostatic equilibration of CO2 supply.Front Plant Sci. 2015 Feb 26;6:106. doi: 10.3389/fpls.2015.00106. eCollection 2015. Front Plant Sci. 2015. PMID: 25767475 Free PMC article.
-
Thylakoid lumen carbonic anhydrase (CAH3) mutation suppresses air-Dier phenotype of LCIB mutant in Chlamydomonas reinhardtii.Plant Physiol. 2009 Feb;149(2):929-37. doi: 10.1104/pp.108.132456. Epub 2008 Dec 12. Plant Physiol. 2009. PMID: 19074623 Free PMC article.
-
The Main Structural and Functional Characteristics of Photosystem-II-Enriched Membranes Isolated from Wild Type and cia3 Mutant Chlamydomonas reinhardtii.Life (Basel). 2020 May 14;10(5):63. doi: 10.3390/life10050063. Life (Basel). 2020. PMID: 32423065 Free PMC article.
References
-
- Badger MR (1985) Photosynthetic oxygen-exchange. Annu Rev Plant Physiol 36: 27-53
-
- Badger MR, Andrews TJ, Whitney SM, Ludwig M, Yellowlees DC, Leggat W, Price GD (1998) The diversity and co-evolution of Rubisco, plastids, pyrenoids and chloroplast-based CCMs in the algae. Can J Bot 76: 1052-1071
-
- Badger MR, Price GD (1994) The role of carbonic anhydrase in photosynthesis. Annu Rev Plant Physiol Plant Mol Biol 45: 369-392
-
- Badger MR, Sharkey TD, von Caemmerer S (1984) The relationship between steady-state gas exchange of bean leaves and the levels of carbon-reduction-cycle intermediates. Planta 160: 305-313 - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
