

Review
doi: 10.1128/jvi.77.17.9109-9115.2003.
Reovirus receptors and pathogenesis
Affiliations
- PMID: 12915527
- PMCID: PMC187431
- DOI: 10.1128/jvi.77.17.9109-9115.2003
Item in Clipboard
Review
Reovirus receptors and pathogenesis
J Virol.
2003 Sep.
No abstract available
Figures
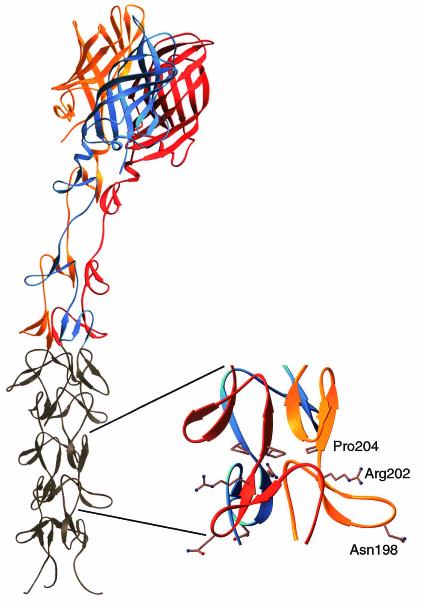
Crystal structure of reovirus attachment protein σ1. The crystal structure of T3D σ1 includes residues 245 to 455 (14). The three monomers of the σ1 trimer are shown in red, orange, and blue. Each monomer consists of a C-terminal head domain formed by a compact β-barrel and an N-terminal fibrous tail that contains three β-spiral repeats. Based on analysis of patterns in aligned σ1 sequences, the β-spiral likely begins at residue 167 of T3D σ1 and comprises eight repeats. The N-terminal five repeats, which are not included in the crystal structure, are shown in gray. The spiral has been extended using translated and rotated σ1 repeats to generate a model that depicts the approximate dimensions of the molecule. Amino acids Asn198, Arg202, and Pro204 have been implicated in the interaction of T3D σ1 with sialic acid (13). The approximate location of these residues in the model (shown in ball-and-stick representation on the right) suggests that they form a binding site for sialic acid. Residues 1 to 167 are not shown; these residues are predicted to form a triple α-helical coiled coil structure (6, 25, 27, 40). This figure was prepared by Thilo Stehle (Harvard University) (published with permission) with the program RIBBONS (10).
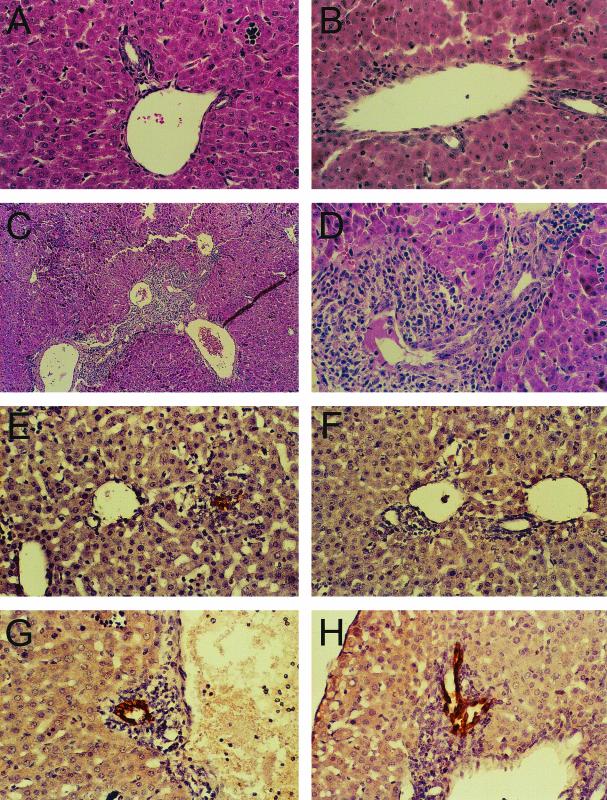
(A to D) Liver histopathology in mice following infection with T3SA− and T3SA+. ND4 Swiss Webster mice (2 to 3 days old) were inoculated perorally with phosphate-buffered saline (A) or 2.5 × 103 PFU of either T3SA− (B) or T3SA+ (C and D). At 8 days postinoculation, liver tissue was harvested, embedded in paraffin, thin sectioned, and stained with hematoxylin and eosin. Magnification, ×100 (C) or ×400 (A, B, and D). (E to H) Immunohistochemical localization of reovirus antigen in bile duct epithelial cells. ND4 Swiss Webster mice (2 to 3 days old) were inoculated perorally with 2.5 × 103 PFU of either T3SA− (E and F) or T3SA+ (G and H). At 6 days postinoculation, liver tissue was harvested, embedded in paraffin, thin sectioned, and stained for reovirus antigen using rabbit anti-reovirus serum and horseradish peroxidase. Dark-brown staining indicates the presence of reovirus antigen. Magnification, ×400. Representative sections from two separate animals are shown. Modified from Barton et al. (5) with permission from the publisher.
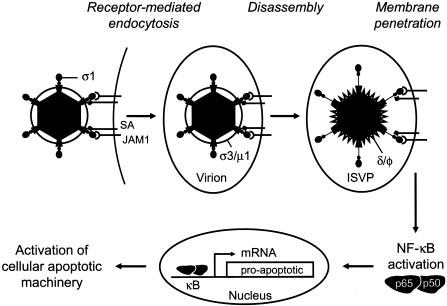
Model of reovirus-induced apoptosis. Reovirus infection is initiated by attachment of the virus to carbohydrate coreceptors and JAM1. For T3 reoviruses, the carbohydrate bound is sialic acid (SA). After attachment to cellular receptors, reovirus enters cells by receptor-mediated endocytosis. Within an endocytic compartment, the viral outer capsid is removed to generate infectious subvirion particles (ISVPs). During virion-to-ISVP conversion, σ3 is degraded and lost from virions, σ1 undergoes a conformational change, and μ1 is cleaved to form particle-associated fragments δ and φ. Removal of σ3 exposes hydrophobic domains in μ1 that facilitate interactions of ISVPs with endosomal membranes, leading to delivery of core particles into the cytoplasm and concomitant activation of the viral transcriptase. Viral attachment and disassembly must occur within the same cellular compartment to activate NF-κB. Activation of NF-κB also might be achieved by μ1-mediated membrane penetration acting in synergy with viral receptor engagement. Once activated, NF-κB translocates to the nucleus, where it induces the expression of proapoptotic genes.
Similar articles
-
Reovirus receptors, cell entry, and proapoptotic signaling.Adv Exp Med Biol. 2013;790:42-71. doi: 10.1007/978-1-4614-7651-1_3. Adv Exp Med Biol. 2013. PMID: 23884585 Free PMC article. Review.
-
The distinct roles of JAM-A in reovirus pathogenesis.Cell Host Microbe. 2009 Jan 22;5(1):3-5. doi: 10.1016/j.chom.2008.12.009. Cell Host Microbe. 2009. PMID: 19154981
-
Structural evidence for common functions and ancestry of the reovirus and adenovirus attachment proteins.Rev Med Virol. 2003 Mar-Apr;13(2):123-32. doi: 10.1002/rmv.379. Rev Med Virol. 2003. PMID: 12627395 Free PMC article. Review.
-
Independent regulation of reovirus membrane penetration and apoptosis by the mu1 phi domain.PLoS Pathog. 2008 Dec;4(12):e1000248. doi: 10.1371/journal.ppat.1000248. Epub 2008 Dec 26. PLoS Pathog. 2008. PMID: 19112493 Free PMC article.
-
Junctional adhesion molecule-A is required for hematogenous dissemination of reovirus.Cell Host Microbe. 2009 Jan 22;5(1):59-71. doi: 10.1016/j.chom.2008.12.001. Cell Host Microbe. 2009. PMID: 19154988 Free PMC article.
Cited by
-
Understanding neurotropic enteric viruses: routes of infection and mechanisms of attenuation.Cell Mol Life Sci. 2024 Oct 4;81(1):413. doi: 10.1007/s00018-024-05450-6. Cell Mol Life Sci. 2024. PMID: 39365457 Free PMC article. Review.
-
Junctional adhesion molecule 1 is a functional receptor for feline calicivirus.J Virol. 2006 May;80(9):4482-90. doi: 10.1128/JVI.80.9.4482-4490.2006. J Virol. 2006. PMID: 16611908 Free PMC article.
-
Manipulation of Host Cell Organelles by Intracellular Pathogens.Int J Mol Sci. 2021 Jun 17;22(12):6484. doi: 10.3390/ijms22126484. Int J Mol Sci. 2021. PMID: 34204285 Free PMC article. Review.
-
Virus-associated disruption of mucosal epithelial tight junctions and its role in viral transmission and spread.Tissue Barriers. 2021 Oct 2;9(4):1943274. doi: 10.1080/21688370.2021.1943274. Epub 2021 Jul 9. Tissue Barriers. 2021. PMID: 34241579 Free PMC article. Review.
-
Virus-Receptor Interactions: The Key to Cellular Invasion.J Mol Biol. 2018 Aug 17;430(17):2590-2611. doi: 10.1016/j.jmb.2018.06.024. Epub 2018 Jun 18. J Mol Biol. 2018. PMID: 29924965 Free PMC article. Review.
References
-
- Azzam-Smoak, K., D. L. Noah, M. J. Stewart, M. A. Blum, and B. Sherry. 2002. Interferon regulatory factor-1, interferon-beta, and reovirus-induced myocarditis. Virology 298:20-29. - PubMed
-
- Banerjea, A. C., K. A. Brechling, C. A. Ray, H. Erikson, D. J. Pickup, and W. K. Joklik. 1988. High-level synthesis of biologically active reovirus protein sigma 1 in a mammalian expression vector system. Virology 167:601-612. - PubMed
-
- Barton, E. S., J. L. Connolly, J. C. Forrest, J. D. Chappell, and T. S. Dermody. 2001. Utilization of sialic acid as a coreceptor enhances reovirus attachment by multistep adhesion strengthening. J. Biol. Chem. 276:2200-2211. - PubMed
-
- Barton, E. S., J. C. Forrest, J. L. Connolly, J. D. Chappell, Y. Liu, F. Schnell, A. Nusrat, C. A. Parkos, and T. S. Dermody. 2001. Junction adhesion molecule is a receptor for reovirus. Cell 104:441-451. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
