

Heterologous human immunodeficiency virus type 1 lentiviral vectors packaging a simian immunodeficiency virus-derived genome display a specific postentry transduction defect in dendritic cells
- PMID: 12915545
- PMCID: PMC187397
- DOI: 10.1128/jvi.77.17.9295-9304.2003
Heterologous human immunodeficiency virus type 1 lentiviral vectors packaging a simian immunodeficiency virus-derived genome display a specific postentry transduction defect in dendritic cells
Abstract
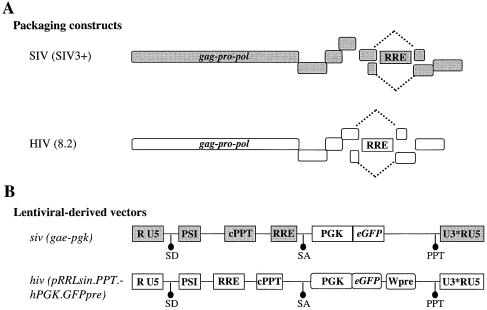
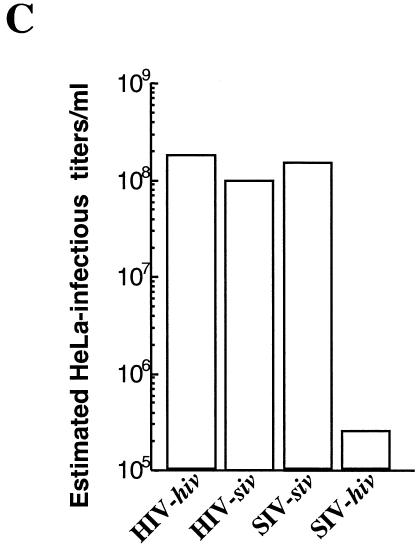
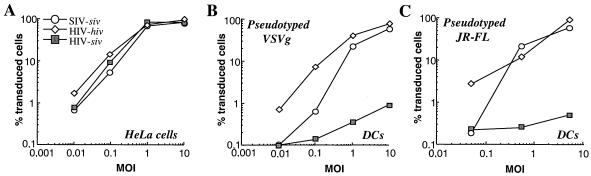
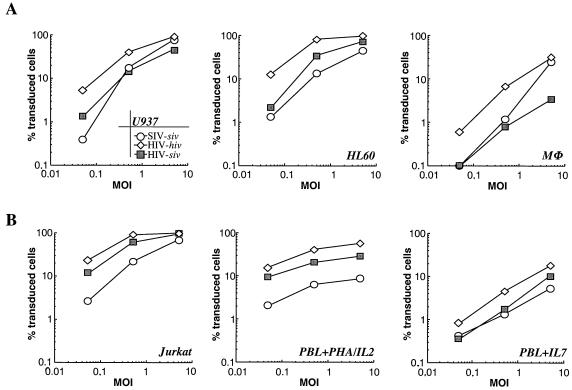
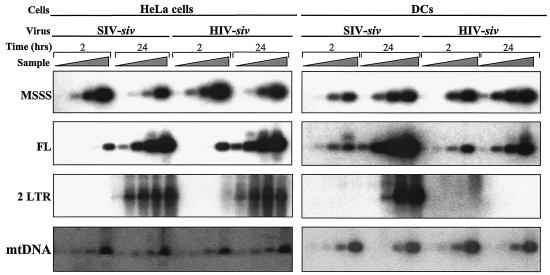
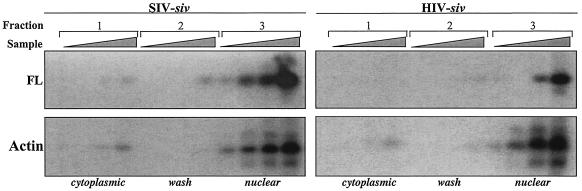
Heterologous lentiviral vectors (LVs) represent a way to address safety concerns in the field of gene therapy by decreasing the possibility of genetic recombination between vector and packaging constructs and the generation of replication-competent viruses. Using described LVs based on human immunodeficiency virus type 1 (HIV-1) and simian immunodeficiency virus MAC251 (SIV(MAC251)), we asked whether heterologous virion particles in which trans-acting factors belonged to HIV-1 and cis elements belonged to SIV(MAC251) (HIV-siv) would behave as parental homologous vectors in all cell types. To our surprise, we found that although the heterologous HIV-siv vector was as infectious as its homologous counterpart in most human cells, it was defective in the transduction of dendritic cells (DCs) and, to a lesser extent, macrophages. In DCs, the main postentry defect was observed in the formation of two-long-terminal-repeat circles, despite the fact that full-length proviral DNA was being synthesized and was associated with the nucleus. Taken together, our data suggest that heterologous HIV-siv vectors display a cell-dependent infectivity defect, most probably at a post-nuclear entry migration step. As homologous HIV and SIV vectors do transduce DCs, we believe that these results underscore the importance of a conserved interaction between cis elements and trans-acting viral factors that is lost or suboptimal in heterologous vectors and essential only in the transduction of certain cell types. For gene therapy purposes, these findings indicate that the cellular tropism of LVs can be modulated not only through the use of distinct envelope proteins or tissue-specific promoters but also through the specific combinatorial use of packaging and transfer vector constructs.
Figures

References
-
- Best, S., P. Le Tissier, G. Towers, and J. P. Stoye. 1996. Positional cloning of the mouse retrovirus restriction gene Fv1. Nature 382:826-829. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
