

Newcastle disease virus V protein is a determinant of host range restriction
- PMID: 12915566
- PMCID: PMC187425
- DOI: 10.1128/jvi.77.17.9522-9532.2003
Newcastle disease virus V protein is a determinant of host range restriction
Abstract
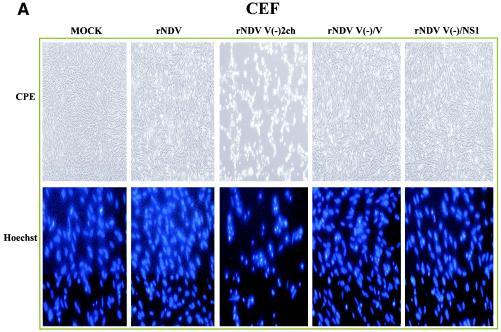
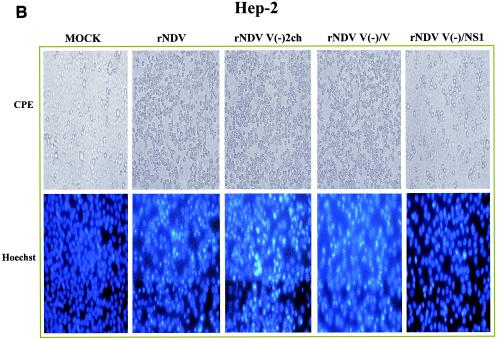
It has been demonstrated that the V protein of Newcastle disease virus (NDV) functions as an alpha/beta interferon (IFN-alpha/beta) antagonist (M. S. Park, M. L. Shaw, J. Muñoz-Jordan, J. F. Cros, T. Nakaya, N. Bouvier, P. Palese, A. García-Sastre, and C. F. Basler, J. Virol. 77:1501-1511, 2003). We now show that the NDV V protein plays an important role in host range restriction. In order to study V functions in vivo, recombinant NDV (rNDV) mutants, defective in the expression of the V protein, were generated. These rNDV mutants grow poorly in both embryonated chicken eggs and chicken embryo fibroblasts (CEFs) compared to the wild-type (wt) rNDV. However, insertion of the NS1 gene of influenza virus A/PR8/34 into the NDV V(-) genome [rNDV V(-)/NS1] restores impaired growth to wt levels in embryonated chicken eggs and CEFs. These data indicate that for viruses infecting avian cells, the NDV V protein and the influenza NS1 protein are functionally interchangeable, even though there are no sequence similarities between the two proteins. Interestingly, in human cells, the titer of wt rNDV is 10 times lower than that of rNDV V(-)/NS1. Correspondingly, the level of IFN secreted by human cells infected with wt rNDV is much higher than that secreted by cells infected with the NS1-expressing rNDV. This suggests that the IFN antagonist activity of the NDV V protein is species specific. Finally, the NDV V protein plays an important role in preventing apoptosis in a species-specific manner. The rNDV defective in V induces apoptotic cell death more rapidly in CEFs than does wt rNDV. Taken together, these data suggest that the host range of NDV is limited by the ability of its V protein to efficiently prevent innate host defenses, such as the IFN response and apoptosis.
Figures
References
-
- Basler, C., A. Reid, J. Dybing, T. Janczewski, T. Fanning, H. Zheng, M. Salvatore, M. Perdue, D. Swayne, A. Garcia-Sastre, P. Palese, and J. Taubenberger. 2001. Sequence of the 1918 pandemic influenza virus nonstructural gene (NS) segment and characterization of recombinant viruses bearing the 1918 NS genes. Proc. Natl. Acad. Sci. USA 98:2746-2751. - PMC - PubMed
-
- Beard, C. W., and R. P. Hanson. 1984. Newcastle disease, p. 452-470. In M. S. Hofstad, H. J. Barnes, B. W. Calnek, W. M. Reid, and W. Yoder (ed.), Disease of poultry. Iowa State University Press, Ames, Iowa.
-
- Garcia-Sastre, A. 2001. Inhibition of interferon-mediated antiviral responses by influenza A viruses and other negative-strand RNA viruses. Virology 279:375-384. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
