

A bromodomain-containing protein from tomato specifically binds potato spindle tuber viroid RNA in vitro and in vivo
- PMID: 12915580
- PMCID: PMC187389
- DOI: 10.1128/jvi.77.17.9685-9694.2003
A bromodomain-containing protein from tomato specifically binds potato spindle tuber viroid RNA in vitro and in vivo
Abstract
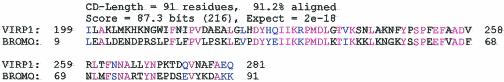
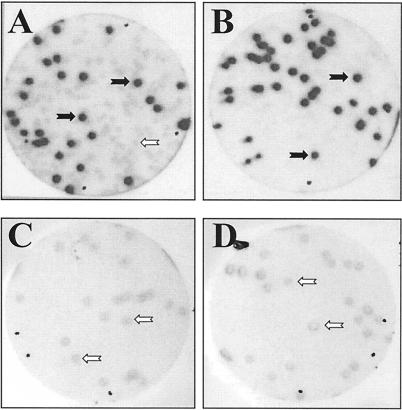
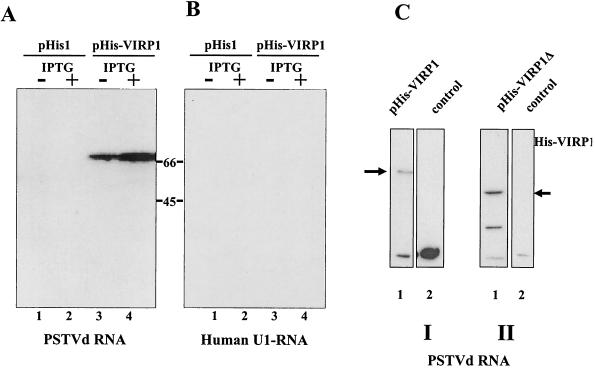
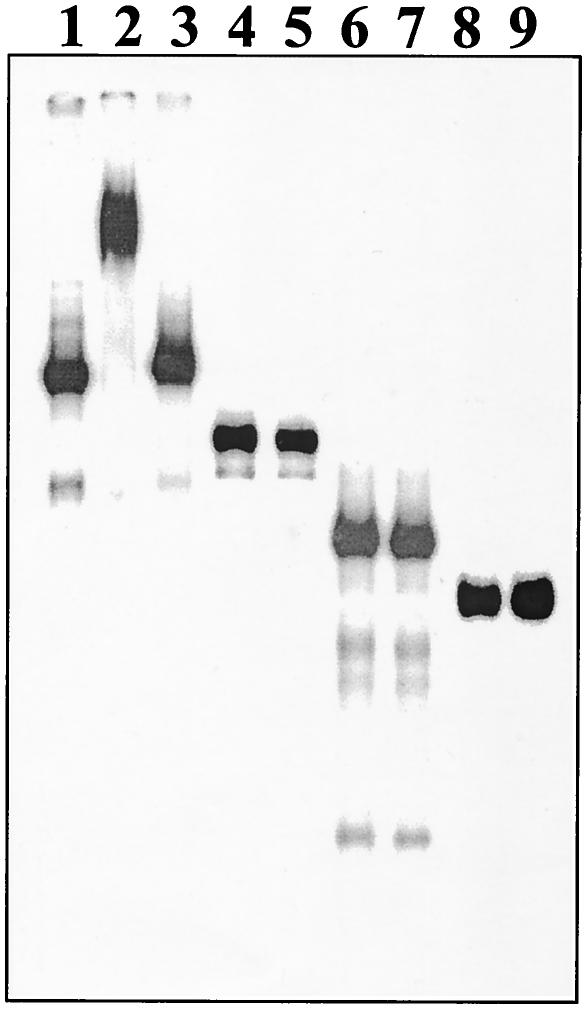
For the identification of RNA-binding proteins that specifically interact with potato spindle tuber viroid (PSTVd), we subjected a tomato cDNA expression library prepared from viroid-infected leaves to an RNA ligand screening procedure. We repeatedly identified cDNA clones that expressed a protein of 602 amino acids. The protein contains a bromodomain and was termed viroid RNA-binding protein 1 (VIRP1). The specificity of interaction of VIRP1 with viroid RNA was studied by different methodologies, which included Northwestern blotting, plaque lift, and electrophoretic mobility shift assays. VIRP1 interacted strongly and specifically with monomeric and oligomeric PSTVd positive-strand RNA transcripts. Other RNAs, for example, U1 RNA, did not bind to VIRP1. Further, we could immunoprecipitate complexes from infected tomato leaves that contained VIRP1 and viroid RNA in vivo. Analysis of the protein sequence revealed that VIRP1 is a member of a newly identified family of transcriptional regulators associated with chromatin remodeling. VIRP1 is the first member of this family of proteins, for which a specific RNA-binding activity is shown. A possible role of VIRP1 in viroid replication and in RNA mediated chromatin remodeling is discussed.
Figures

References
-
- Barlev, N. A., V. Poltoratsky, T. Owen-Hughes, C. Ying, L. Liu, J. L. Workman, and S. L. Berger. 1998. Repression of GCN5 histone acetyltransferase activity via bromodomain-mediated binding and phosphorylation by the Ku-DNA-dependent protein kinase complex. Mol. Cell. Biol. 18:1349-1358. - PMC - PubMed
-
- Beck, S., I. Hanson, A. Kelly, D. J. Pappin, and J. Trowsdale. 1992. A homologue of the Drosophila female sterile homeotic (fsh) gene in the class II region of the human MHC. DNA Seq. 2:203-210. - PubMed
-
- Chamberlin, H. M., and J. H. Thomas. 2000. The bromodomain protein LIN-49 and trithorax-related protein LIN-59 affect development and gene expression in Caenorhabditis elegans. Development 127:713-723. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Research Materials
